Lindens make great trees for urban areas. A few species and hybrids in particular are commonly planted in parks, yards, and along the streets of cities across the northern hemisphere and have been for decades – centuries even. They cast dense shade, are tolerant of a variety of climates and soil conditions, and are generally easy to maintain. For much of the year as you move throughout the city you live in, you likely pass by dozens of lindens without thinking twice about them. They are ubiquitous, conventional, ordinary, common. Unless they’re in bloom. For a few weeks in early to mid-summer, flowering lindens produce an impossibly sweet fragrance that can’t be ignored. Along with the scent comes the sound of hundreds of buzzing bees collecting pollen and nectar from the pendulous blooms.
Lindens are trees and shrubs in the family Malvaceae and genus Tilia. Around 30 or so species are found in temperate regions across the northern hemisphere, mostly in Europe and Asia. Depending on who you ask, there are between one and three species native to North America. Tilia caroliniana and Tilia heterophylla are considered by some to be varieties of Tilia americana, or American basswood, which is distributed across central and eastern United States and north into parts of Canada. Another common name for linden is lime because words used to refer to the tree in older languages were similar to the word lime. The name basswood comes from the tree’s fibrous inner bark, known as bast.

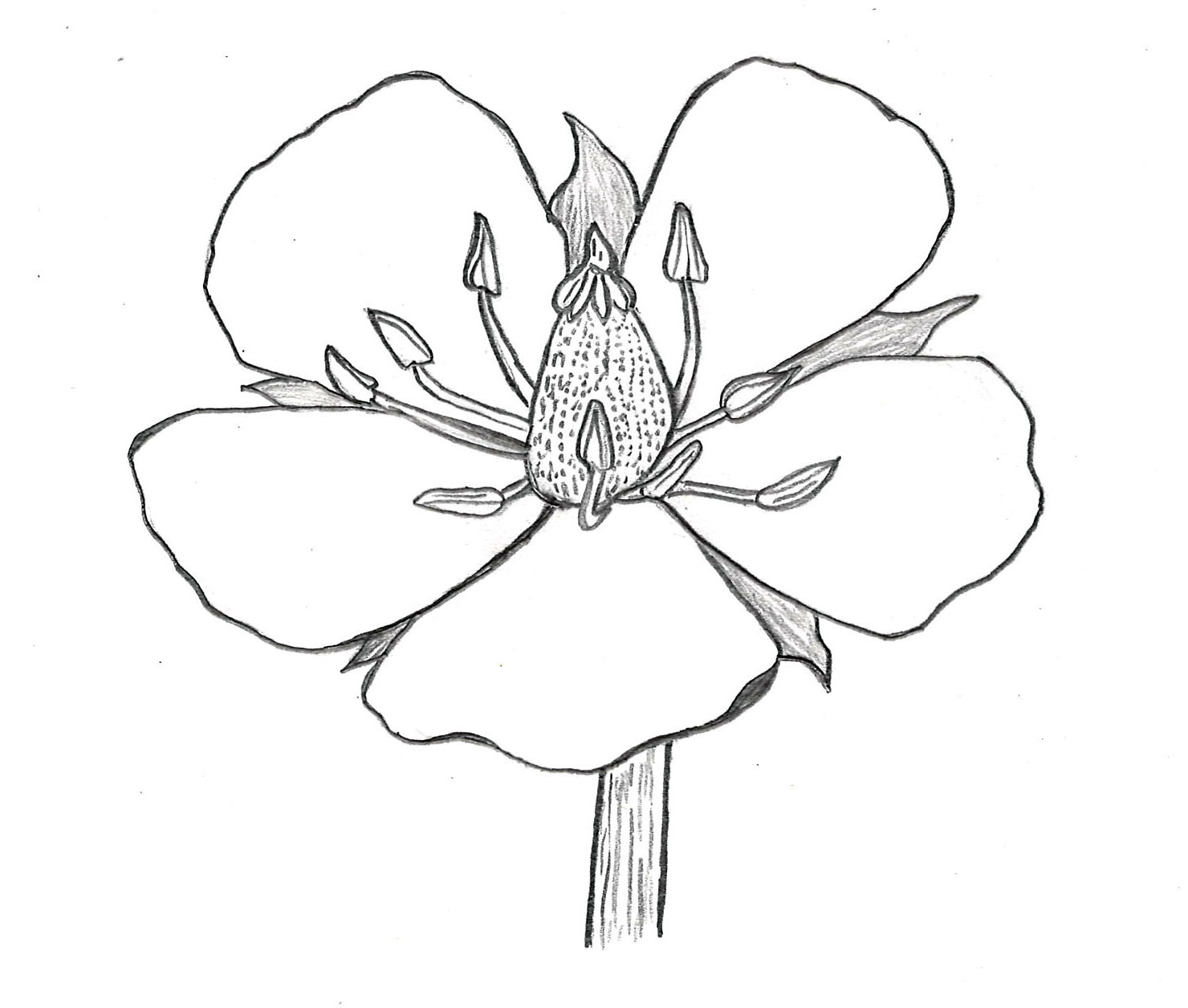
Linden leaves are generally heart-shaped and asymmetrical with serrate margins. Small clusters of little yellow to white flowers form at the end of a slender stem attached to a narrow, ribbon-like, yellow-green bract. The bract aids in seed dispersal by helping the fruits float on the wind away from the parent tree in a manner similar to the samaras of maple trees. The fruits are small, round, hardened drupes that resemble little peas. The fragrant, nectar-rich flowers are not only favored by beekeepers for honey production, but also have a long history of being harvested for making tea (i.e. tisane). Linden flower tea is said to have a number of medicinal uses and health benefits, all of which I take with a grain of salt. This series of posts isn’t meant to be an investigation into the health claims of plants, but instead an opportunity for me – out of sheer curiosity – to try making tea out of a variety of different plants . If medicinal uses interest you, I encourage you to seek out credible, peer-reviewed sources.
I made linden flower tea from flowers I collected from Tilia cordata, commonly known as littleleaf linden. It was an easy one to find due to its popularity as an urban tree. The natural distribution of littleleaf linden extends from Britain across Europe and into western Asia. Its triangular-ovate shaped leaves are 4-10 centimeters long, glossy green on top, and pale green on the bottom with tufts of orange hairs along the leaf veins, concentrated at the base of the leaf where the leaf blade meets the petiole. The tree can reach up to 21 meters tall and has an oval or rounded-pyramidal shape, though many trees in urban areas are cultivars and can be smaller and more compact.

I harvested the flowers – bracts and all – in late June. It’s advised that they not be harvested directly after a rain (or after being hit by sprinklers), and that they are harvested when the flowers are newly opened. I presume this is because the flowers are at their freshest at this point and will be the best for making tea. I layed the flowers out to dry on a clean kitchen towel on top of a metal cake rack. It only takes 2 or 3 days for them to dry. After drying I removed and saved all the flowers and threw out the bracts and stems, but apparently you can use the entire inflorescence if you’d like.
There are several linden flower tea recipes online. I went with 3 cups of boiling water poured over 1 tablespoon dried linden flowers, covered and steeped for 15 minutes. The resulting tea was an appealing pastel yellow color. I tried it plain as well as sweetened with a little bit of honey. I preferred it sweetened, but unsweetened wasn’t too bad, just a little bitter. It has a floral taste and pleasant smell. Sierra said it tasted earthy, like something she wasn’t supposed to be drinking. Despite that odd review, she said she liked it. Since several sources discussed the calming, sleep-inducing effects of the tea, I made sure to drink it in the evening when it would be normal for me to be feeling sleepy. I suggest you do the same.