My book is out! It showed up a week earlier than expected, a pleasant surprise for anyone who pre-ordered a copy. Common Backyard Weeds of the Pacific Northwest is available now from your favorite bookseller. You can also order it directly fromthe publisher or through my Bookshop store. Thank you for your support! If and when you get your hands on a copy, please tell me what you think. Constructive feedback would be particularly helpful so that I can make improvements to the second edition. (Yes, I’m already planning on a second edition, and I already have a few edits of my own!)
While researching this book, I came across many new and interesting facts about weeds that I didn’t previously know. It’s part of the joy of writing. Some of these discoveries pertain to cocklebur, a prickly and troubling weed with a fascinating dispersal story. Xanthium strumarium is a widespread plant that comes up in farms, rangelands, wetlands, yards, vacant lots, roadsides, ditch banks, and a variety of other locations. In the book I introduce it this way: “A native and/or introduced annual (short-lived perennial) that grows up to 4 feet tall with either a single or a few rigid stems.” It produces a thick, woody taproot and has broad, simple leaves. Separate male and female flowers occur on the same plant, and the flowers are wind-pollinated and predominantly self-pollinated – a unique state of affairs for a plant in the aster family.
fruits of cocklebur (Xanthium strumarium)
The most recognizable feature of cocklebur are the fruits, which are egg-shaped and covered in hooked prickles. One of their dispersal mechanisms is immediately obvious, grabbing onto passers-by and hitching a ride. Since they are often found near waterways or in areas where flooding occurs, another way they are dispersed is by floating in water. Both of these dispersal pathways allow seeds to travel long distances.
Still, cockleburs have another trick up their sleeves that helps ensure their longevity. Inside each fruit there are two seeds which look a bit like oversized sunflower seeds. These seeds exhibit something called somatic polymorphism. This term can mean different things depending on the plant, but in cocklebur’s case it means that the two seeds have different dormancies. One seed will germinate within the first year. The second seed remains dormant for at least a year and can germinate much later. A classic example of bet hedging. The two seeds are equipped with a water-soluble germination inhibitor. Oxygen degrades the inhibitors which leads to germination, but this happens at a different rate in each of the two seeds. (Read more about this here.)
inside the fruit of a cocklebur
From a weed management perspective, dispersal mechanisms and seed viability are two critical traits to become familiar with. When you know how your weeds get around and how long their seeds might last in the soil, you can make informed decisions on which weeds to remove and when. Just another reason to get my book!
_____________________
If you are in the Boise area, I will be at Rediscovered Books this Saturday (July 11,2026) for a short presentation and book signing. Goherefor more details. Follow me on Instagram to stay informed on this and future book events. Hope to see you there!
Sweetgum is a distinctive tree at any time of year. It stands out among most other deciduous trees with its excurrent growth habit, which gives it a narrowly conical or pyramidal shape. Only in its advanced age does it begin to develop a more rounded and irregular form. Its leaves are star-shaped with either five or seven lobes and span between four and seven inches. Their glossy green color gives way to a wide range of colors in the fall, including yellow, orange, red, and maroon, remaining on the tree for several weeks depending on weather conditions. The fruits are particularly distinct, especially in the winter after the leaves have dropped. Woody balls made up of a series of capsules with pointed beaks hang from long stalks throughout the winter, windborne seeds having been released from small openings in the fall. Without even bothering to look at the twigs, seeing these durable, spiky balls hanging from branches (or feeling them under your feet) is a quick indication that you are looking at a sweetgum.
one of manyfall colors found on sweetgum (Liquidambar styraciflua)
Liquidamabar styraciflua (one of the funnest botanical names to say) was previously placed in the family Hamamelidaceae but is now one of the few members of Altingiaceae. Its natural distribution is broad, covering a large portion of the eastern United States and west into Missouri then down into Texas, Mexico, and much of Central America. Outside of its natural range it has been widely planted as an ornamental, and there are several popular sweetgum cultivars currently in cultivation. Both the common and botanical names for this tree refer to the resin found in its bark, which historically has had many uses.
The winter twigs of sweetgum are stout, round, smooth, and yellow-brown to green or olive-green to brown-purple in color. They can also be glossy and feature a few scattered lenticels. Older twigs (or branchlets) are brown at first and then light grey with dark grey lenticels. They are occasionally adorned with corky wings similar to those of bur oak. Because the wings aren’t always present, it can be a fun thing to encounter when you are out looking at twigs.
corky wings on the branchlets of sweetgum
The winter buds of sweetgum twigs are egg-shaped and made up of bud scales with acute tips and ciliate margins. They are green to orange-brown or reddish in color and occasionally sticky. Lateral buds are alternately arranged, are much smaller than terminal buds, and are stalkless and flattened against the twig. They sit above a slightly raised leaf scar that is half-elliptical to triangular in shape and has three distinct vascular bundle traces. The pith of sweetgum twigs is solid, continuous, brownish, and irregularly shaped.
winter twig of sweetgum tree
Sweetgum bark is light to dark grey and is made up of a series of rough, vertically arranged, scaly ridges that become deeply furrowed with age. The mace-like fruits of sweetgum are about one inch wide and, at a glance, are similar in appearance to the seed balls of sycamore trees; however, sycamore seed balls easily break apart when compressed, while the pointed, woody capsules that make up a sweetgum ball are held firmly together and can hold their shape for long periods of time. When these “gumballs” collect on the ground below, they can become a hazard, especially where there is lots of foot traffic. Speaking from experience, they are also obnoxious when operating a mower. This polarizing feature has resulted in bad opinions of the sweetgum tree. Luckily, some people are out there defending it.
lf you’ve seen one fireweed, you’ve probably seen several. As an early successional species, growing in large numbers across a vast amount of space is kind of its thing. Any disturbance that leaves bare ground in its wake, such as a wildfire or a windstorm, gives fireweed the opportunity to colonize. It grows quickly and spreads via rhizomes, producing thousands of airborne seeds in the process, sending them off to continue colonization or contribute to soil seed banks in preparation for future disturbances. The role of plants like fireweed is vital – promptly covering bare ground to stave off erosion and acting as a nurse plant to new saplings destined to become the future forest. In a garden setting or in locations where aggressively spreading plants are discouraged, fireweed and its weedy behavior may be unwelcome, but in other contexts, its services are essential.
fireweed (Chamerion angustifolium)
Fireweed (Chamerion angustifolium) is a species in Onagraceae, commonly known as the evening primrose family. It has an impressive distribution, widespread across much of North America and Eurasia. This is owed largely to its adaptability. Deep shade and overly dry soil are two conditions that it does not tolerate well, otherwise it seems to grow in a wide variety of soil types, moisture levels, and sun exposures, particularly in areas where there is regular disturbance. Swaths of towering plants topped with rose-pink flower spikes make fireweed impossible to ignore and a favorite of wildflower enthusiasts.
Fireweed stems reach from three to nine feet tall and are rarely branched. Long, narrow, lance-shaped leaves are arranged alternately along the lengths of stems and give the plant a willow-like appearance, which explains another common name, rosebay willowherb. The undersides of leaves have a distinct venation pattern, in which the veins don’t reach the leaf margins, a feature that can help with identification.
distinct leaf veins of fireweed(Chamerion angustifolium)
A series of rose-pink to purple flowers top the stems of fireweed. Each flower has four sepals and four petals with eight stamens and a four-lobed stigma extending prominently from the center. Its long, narrow ovary can be confused for a flower stalk. Rich with nectar, fireweed flowers are a favorite of honeybee keepers. They are also edible, like much of the rest of the plant. Narrow, four-chambered capsules form in place of fertilized flowers and later split open to release abundant, small seeds with a tuft of fluff attached to each one to aid in wind dispersal.
Fireweed has a long history of being used as food and medicine. Stem fibers are also useful for making cord, and seed fluff is useful for weaving and padding. Certainly, fireweed’s abundance and ubiquity contribute to its utility. Having never eaten fireweed before, I decided that a good way to introduce it to my diet would be to make a tea. Fireweed leaves are commonly collected for tea and are said to make an excellent non-caffeinated replacement for black tea.
fireweed tealeaves
Making fireweed tea starts by stripping young leaves from fireweed stems. Recipes I encountered all called for fermenting the leaves before drying them. I did this by squeezing handfuls of leaves in my fists just enough to break and bruise them a bit and then packing them into a quart size Mason jar. I closed the lid tight and kept them in the jar for about five days, shaking it up a couple times a day supposedly to help prevent mold issues. After that, I dried the leaves on a baking sheet in the hot sun. From there, they are ready for making tea the same way you would make any other loose leaf tea, chopping the leaves up a bit before immersing them in hot water.
I found the taste of fireweed tea to be mild and pleasant. Despite several sources comparing it to black tea, I thought it was more similar to green tea. Sierra liked the smell more than the taste and wished it had honey in it. Compared to other teas I’ve tried in this short series of posts, this is definitely one of the better ones, and a tea I could see myself making again sometime.
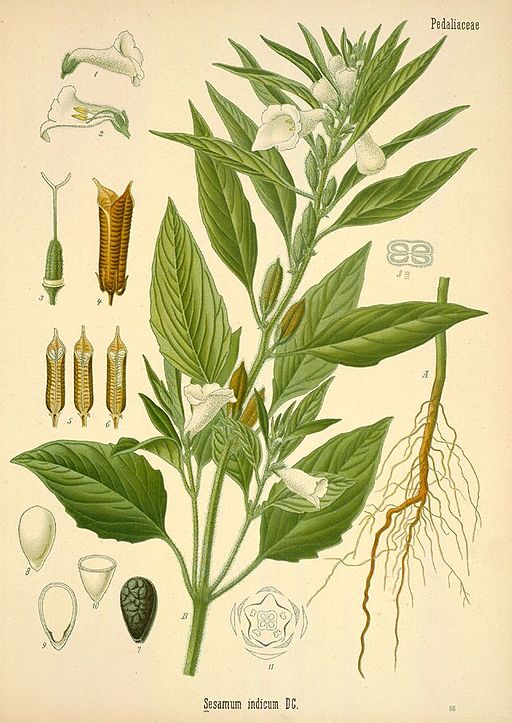
In certain instances, “open sesame” might be something you exclaim to magically open the door to a cave full of treasure, but for the sesame plant, open sesame is a way of life. In sesame’s case, seeds are the treasure, which are kept inside a four-chambered capsule. In order for the next generation of plants to have a chance at life, the seeds must be set free. Sesame’s story is similar to the stories of numerous other plant species whose seeds are born in dehiscent fruits. But in this instance, the process of opening those fruits is fairly unique.
Sesamum indicum is a domesticated plant with a 5000 plus year history of cultivation. It shares a genus with about 20 other species – most of which occur in sub-Saharan Africa – and belongs to the family Pedaliaceae – the sesame family. Sesame was first domesticated in India and is now grown in many other parts of the world. It is an annual plant that is drought and heat-tolerant and can be grown in poor soils and locations where many other crops might struggle. However, the best yields are achieved on farms with fertile soils and adequate moisture.
Depending on the variety and growing conditions, sesame can reach up to 5 feet tall and can be unbranched or highly branched. Its broad lance-shaped leaves are generally arranged directly across from each other on the stem. The flowers are tubular, similar in appearance to foxglove, and are typically self-pollinated and short-lived. They come in shades of white, pink, blue, and purple and continue to open throughout the growing season as the plant grows taller, even as fruits formed earlier mature. The fruits are deeply-grooved capsules with at least four separate chambers called locules. Rows of tiny, flat, teardrop shaped seeds are produced in each chamber. The seeds are prized for their high oil content and are also used in numerous other ways, both processed and fresh. One of my favorite uses for sesame seeds is tahini, which is one of the main ingredients in hummus.
The fruits of sesame are dehiscent, which means they naturally split open upon reaching maturity. Compare this to indehiscent fruits like acorns, which must either rot or be chewed open by an animal in order to free the seeds. Dehiscence is also called shattering, and in many domesticated crop plants, shattering is something that humans have selected against. If fruits dehisce before they can be harvested, seeds fall to the ground and are lost. Selecting varieties that hold on to their seed long enough to be harvested was imperative for crops like beans, peas, and grains. In domesticated sesame, the shattering trait persists and yield losses are often high.
Most of the world’s sesame crop is harvested by hand. The plants are cut, tied into bundles, and left to dry. Once dry, they are held upside down and beaten in order to collect the seeds from their dehisced capsules. When harvested this way, naturally shattering capsules may be preferred. But in places like the United States and Australia, where mechanical harvesting is desired, it has been necessary to develop new, indehiscent varieties that can be harvested using a combine without losing all the seed in the process. Developing varieties with shatter-resistant seed pods, has been challenging. In early trials, seed pods were too tough and passed through threshers without opening. Additional threshing damaged the seeds and caused the harvest to go rancid. Mechanically harvested varieties of sesame exist today, and improvements in these non-shattering varieties continue to be made.
In order to develop these new varieties, breeders have had to gain an understanding of the mechanisms behind dehiscence and the genes involved in this process. This research has helped us appreciate the unique way that the capsules of the sesame plant dehisce. As in the seed bearing parts of many other plant species, the capsules of sesame exhibit hygroscopic movements. That is, their movements are driven by changes in humidity. The simplest form of hygroscopic movement is bending, which can be seen in the opening and closing of pine cone scales. A more complex movement can be seen in the seed pods of many species in the pea family, which both bend and twist as they split open. In both of these examples, water is evaporating from the plant part in question. As it dries it bends and/or twists, thereby releasing its contents.
The cylindrical nature and cellular composition of sesame fruits leads to an even more complex form of hygroscopic movement. Initially, the capsule splits at the top, creating an opening to each of the four locules. The walls of each locule bend outward, then split and twist as the seed falls from the capsule. In a study published in Frontiers in Plant Science (2016), researchers found that differences in the capsule’s inner endocarp layer and outer mesocarp layer are what help lead to this interesting movement. The endocarp layer is composed of both transvere (i.e. circumferential) and longitudinal fiber cells, while the mesocarp is made up of soft parenchyma cells. The thicknesses of these two layers gradually changes along the length of the capsule. As the mesocarp dries, the capsule initially splits open and starts bending outwards, but as it does, resistance from the fiber cells in the endocarp layer causes further bending and twisting (see Figure 1 in the report for an illustration). As the researchers write, “the non-uniform relative thickness of the layers promotes a graded bi-axial bending, leading to the complex capsule opening movement.”
All this considered, a rock rolling away from the entrance of a cave after giving the command, “Open sesame!” almost seems simpler than the “open sesame” experienced by the fruit of the sesame plant.
Probably the most well known strategy that plants have for dispersal is by way of seeds. Seeds are plants in embryo, and new generations of plants are born when seeds, released from their parent plants, find suitable locations to germinate. But one of the most amazing things about plants in general is that they have the ability to reproduce in a variety of different ways, and many plant species are not limited to seeds as their only means of dispersal. A paper by Scott Zona and Cody Coyotee Howard, published in Flora (February 2022), introduces us to the intriguing world of aerial vegetative diaspores – just one of the many ways that plants have to get around.
A diaspore is a plant structure that facilitates dispersal. Seeds are diaspores, as are spores, which are produced by non-seed bearing plants like mosses and ferns. If you’ve ever planted bulbs, you’ve handled another type of diaspore. Bulbs and corms, which many spring flowering plants are grown from, form little offshoots called bulblets and cormels that, when detached from their parent structure, can grow into new individuals. These vegetative diaspores are produced below ground. Aerial vegetative diaspores, on the other hand, are formed on above ground plant parts. This clunky term encompasses a number of different structures that are often simply called bulbils, which Zona and Howard explain is used as “a catch-all term that obscures their morphological identity.”
Compiling a list of plant species that feature aerial vegetative diaspores is a difficult task when plant descriptions from various sources use a broad selection of terminology for the same or similar plant parts. To help complete this task, Zona and Howard defined five distinct types of aerial vegetative diaspores – plantlets, bulbils, cormlets, tubercles, and gemmae – and came up with a list of 252 taxa that are known to feature at least one of these structures.
Plantlets are miniature plants attached to another plant. Once mature, they have clearly visible leaves, stems, and roots (or root initials) and are non-dormant, meaning they are ready to grow on their own as soon as they’re given the opportunity. The tiny plants borne along the margins of the leaves of mother of thousands (Kalanchoe daigremontiana) is a great example of a plantlet.
A bulbil consists of a shortened stem surrounded by scale leaves modified for food and water storage. Sometimes root initials are visible at the base of the bulbil. Bulbils remain dormant until they are dispersed and conditions are suitable for growth. When bulbils start growing but remain attached to the plant, they become a plantlet. A good example of a bulbil can be found on bulbous bluegrass (Poa bulbosa).
Cormlets are comprised of stem tissue and, like plantlets and bulbils, have a single axis of polarity. They have highly reduced scale leaves and are dormant at dispersal. Bulbil bugle lily (Watsonia meriana), despite its misleading common name, is a good example of a plant that produces cormlets.
Tubercles are made up of swollen stem tissue and, like tubers (their underground counterparts), have multiple shoot buds and multiple axes of polarity (meaning there is no right side up like there is in plantlets, bulbils, and cormlets). They lack scale leaves and are dormant at dispersal. Air potato (Dioscorea bulbifera) is an example of a tubercle-producing plant. As you might guess from the common name, potato-like structures are produced aerially on this vining plant that was introduced to North America from Africa and is now invasive in Florida.
A gemma is a tiny cluster of undifferentiated cells. Gemmae are non-dormant and lack polarity. They are the smallest and least common form of aerial vegetative diaspore and can be found on Drosera pygmaea, a species of sundew native to parts of Australia and New Zealand.
Zona and Howard’s list of plants with aerial vegetative diaspores is the most comprehensive list to date – although it is undoubtedly and understandably missing some – and includes representatives from 42 plant families and 21 plant orders. Plantlets are the most common form of aerial vegetative diaspore at 116 taxa, with bulbils coming in second at 72. Cormlets and tubercles are less common, with 25 and 16 taxa respectively. Their paper includes the full list and offers further information about many of the species listed. It’s worth taking time to explore and is a valuable resource for anyone interested in the topic. In addition, their discussion section highlights a number of questions that warrant further investigation.
Questions surrounding reproductive strategies and the dispersal of aerial vegetative diaspores are particularly interesting. Because these structures are vegetative, they are essentially clones of the parent plant, meaning there is no genetic mixing as occurs when seeds are produced. This can be an advantage when sexual reproduction isn’t possible due to lack of pollinators, environmental restrictions, or chromosomal/polyploidy anomalies. It also assures that new individuals are pre-adapted to the site, and it can help a species colonize an area quickly. This ability to rapidly colonize explains why several of the species on Kona and Howard’s list are known to be invasive in parts of the world outside of their native range.
A species that produces both seeds and aerial vegetative diaspores may have an advantage when it comes to dispersal since both types of diaspores have their strengths. Seeds can remain dormant in the soil and are likely to persist in the environment longer than vegetative diaspores, but vegetative diaspores can be produced without relying on pollinators and can establish new individuals quickly. The modes of dispersal between the two can also vary. Seeds can be dispersed by wind or carried away by animals, while vegetative diaspores often rely solely on gravity to get around. One exception is hitchhiker elephant ear (Remusatia vivipara), whose bulbils are equipped with tiny hooks that cling to animal fur and are transported in a similar manner to burs.
When the advantages of aerial vegetative diaspores are considered, it is a wonder that we don’t see them more often. Many plants can be easily propagated by taking stem, leaf, and/or root cuttings and placing them in conditions that favor adventitious root and shoot growth. This may suggest that dormant genetic pathways for producing vegetative diaspores exist in most plants. Or maybe not. Genetic studies of species that feature these structures are needed in order to understand why they are found in some species and not others. Kona and Howard leave us with a slew of research questions like this, and it’s a topic I’ll continue to check in on.
It is said that the inspiration for Velcro came when Swiss inventor, George de Mestral, was removing the burrs of burdock from his dog’s coat, an experience we had with Kōura just days after adopting her. I knew that common burdock was found on our property, and I had made a point to remove all the plants that I could easily get to. However, during Kōura’s thorough exploration of our yard, she managed to find the one plant I had yet to pull due to its awkward location behind the chicken coop.
I knew when I saw the clump of burrs attached to her hind end that we were going to spend the evening combing them out of her fur. However, not long after that we discovered that Kōura had already started the process and in doing so had either swallowed or inhaled some. What tipped us off was her violent hacking and gagging as she moved frantically around the living room. She was clearly distraught, and so were we. Recognizing that she had probably swallowed a burr, we made a quick decision to take her to an emergency vet. This was our unfortunately timed (this happened on Christmas Eve) introduction to burr tongue and all the frightening things that can happen when a dog swallows burdock burrs.
The roots, shoots, and leaves of both greater burdock (Arctium lappa) and common burdock (Arctiumminus) are edible, which I have already discussed in an Eating Weeds post. The burrs, on the other hand, are clearly not. While sticking to the fur of animals and the clothing of people is an excellent way for a plant to get their seeds dispersed, the sharp, hooked barbs that facilitate this are not something you want down your throat. When this occurs, the natural response is to try to hack them up, which Kōura was doing. Salivating heavily and vomiting can also help. In many cases, this will be enough to eliminate the barbs. However, if they manage to work their way into the soft tissues of the mouth, tongue, tonsils, or throat and remain there, serious infection can occur.
burr of common burdock (Arctium minus)
A paper published in The Canadian Veterinary Journal in 1973 describes the treatment for what is commonly known as burr tongue and technically referred to as granular stomatitis. The paper gives an account of what can happen when “long-haired breeds of dogs … run free in areas where [burdock] grows” and the hooked scales of the burrs consequently “penetrate the mucous membrane of the mouth and tongue.” Dogs with burrs imbedded in their mouths may start eating less or more slowly, drinking more water, and drooling excessively. As infection progresses, their breath can start to stink. A look inside the mouth and at the tongue will reveal lesions where the burrs have embedded themselves. Treatment involves putting the dog under anesthesia, scraping away the infected tissue, and administering antibiotics. Depending on the severity of the lesions, scar tissue can form where the barbs were attached.
To prevent infection from happening in the first place, a veterinarian can put the dog under anesthesia and use a camera inside the dog’s mouth and throat to search for pieces of the burr that may have gotten lodged. There is no guarantee that they will find them all or be able to remove them, and so the dog should be monitored over the next several days for signs and symptoms. At our veterinary visit, the vet also warned us that if any burrs were inhaled into the lungs, they could cause a lung infection, which is another thing to monitor for since it would be practically impossible for an x-ray or a camera to initially find them.
Luckily, now more than three weeks later, Kōura appears to be doing fine, and the offending burdock has been taken care of. One thing is for sure, as someone who is generally forgiving of weeds, burdock is one weed that will not be permitted to grow at Awkward Botany Headquarters.
For more adventures involving Kōura, be sure to follow her on Instagram @plantdoctordog.
Behind the scales of a pine cone lie the seeds that promise future generations of pine trees. Even though the seeds are not housed within fruits as they are in angiosperms (i.e. flowering plants), the tough scales of pine cones help protect the developing seeds and keep them secure until the time comes for dispersal. In some species, scales open on their own as the cone matures, at which point winged seeds fall from the tree, taking flight towards their new homes. In other species, the scales must be pried open by an animal in order to free the seed. A third group of species have what are called serotinous cones, the scales of which are sealed shut with resin. High temperatures are required to soften the resin and expose the seeds.
Serotinous cones are a common trait of pine species located in regions where wildfire naturally and regularly occurs. One such species is lodgepole pine (Pinus contorta), which is found in abundance in forests across much of western North America. Lodgepole pine is a thin-barked tree species that burns easily and is often one of the first plants to recolonize after a stand-replacing wildfire. There are 3 or 4 subspecies of lodgepole pine. The one with the largest distribution and the one that most commonly exhibits serotinous cones is P. contorta subsp. latifolia, which occurs throughout the Rocky Mountains, north into the Yukon, and just west of the Cascade Range.
needles of lodgpole pine (Pinus contorta)
Lodgepole pine grows tall and straight, generally maxing out at around 80 feet tall. Its needles are about two and a half inches long, are borne in bundles of two, and tend to twist away from each other, which is one explanation for the specific epithet, contorta. Its cones are egg-shaped with asymmetrical bases, measuring less than two inches long with prickly tips at the ends of each scale. The seeds of lodgepole pine are tiny with little, papery wings that aid in dispersal. The cones can remain attached to the tree for 15-20 years (sometimes much longer), and the seeds remain viable for decades. In non-serotinous cones, the scales start opening on their own in early autumn. Serotinous cones require temperatures of 45-50°C (113-122°F), to release the resin bond between the scales. Some cones that happen to fall from the tree can open when exposed to particularly warm temperatures on the ground. Otherwise, it takes fire to free the seeds.
Serotinous cones aren’t a guarantee, and the percentage of trees with serotinous cones compared to those with non-serotinous cones varies widely across the range of lodgepole pine, both in space and in time. One reason for this is that trees with serotinous cones don’t develop them until they reach a certain age, generally around 20-30 years old, or perhaps as old as 50 or 60. The cones of young trees are all non-serotinous. But some trees never develop serotinous cones at all. Serotiny is a genetic trait, and there are various factors that either select for or against it. A number of factors are at play simultaneously over the life of a tree and across a population of trees, so it is difficult to determine exactly why the percentage of serotinous cones is so variable across the range of the species. What follows are a few potential explanations for this phenomenon.
closed cone of lodgepole pine (Pinus contorta)
As a fire-adapted, pioneer species, lodgepole pine has evolved to live in environments where fire is predictably common. Serotinous cones help ensure that a population won’t be wiped out when a massive wildfire comes through. After the fire has passed and the seeds are released, lodgepole pine can quickly repopulate the barren ground. As long as fire occurs within the lifespan of a population of similarly aged trees, it is advantageous for the majority of individuals to maintain their serotinous trait. If the population is located in an area that historically does not see much fire, serotinous cones may be a disadvantage and can have adverse effects on the longevity of that population.
A study published in Ecology in 2003looked at the influence that the frequency of fire has on lodgepole pine stands found at low and high elevations in Yellowstone National Park. At lower elevations, where summer temperatures are warmer and precipitation is relatively minimal, fires occur more frequently compared to higher elevations, which tend to be cooler and wetter. The researchers found that at lower elevations when fires occurred at short intervals (less than 100 years between each fire), lodgepole pine was slower to repopulate compared to longer intervals. This suggests that the percentage of serotiny found in stands that experienced short fire intervals was low, and that stands with long fire intervals exhibit a higher percentage of serotiny. After all, as mentioned above, lodgepole pines don’t start developing serotinous cones until later in life.
At higher elevations, where fire occurs less frequently, lodgepole pines were found to have a low percentage of serotinous cones regardless of the age of the stand. Because the trees at high elevations are more likely to die of old age rather than fire, maintaining serotinous cones would be a disadvantage. Open cones are preferred. Thus, at least in this study, a greater percentage of serotinous cones was found in lodgepole pines at lower elevations compared to those at higher elevations. Latitude, elevation, mountain pine beetle attacks, and other environmental factors have all been used to explain differences in serotiny. However, the factor that seems to have the greatest influence is the frequency of fire. As James Lotan writes in a 1976 report: “A high degree of cone serotiny would be expected where repeated, high-intensity fires occur. Where forest canopies are disrupted by factors other than fire, open cones annually supply [seed] for restocking disturbances such as windfalls.”
That being said, one other factor does appear to play a critical role in whether or not lodgepole pines produce serotinous cones, and that is seed predation by squirrels. In a paper published in Ecology in 2004, researchers wondered why the percentage of serotinous cones wasn’t even higher in populations where fire reliably occurred during the lifetime of the stand. To help answer this question they looked at the activities of pine squirrels, which are the main seed predator of lodgepole pine seeds. Pine squirrels visit the canopy of lodgepole pines and consume the seeds found in serotinous cones. Because non-serotinous cones quickly shed their seeds, serotinous cones are a more reliable and accessible food source, and because pine squirrels are so effective at harvesting the seeds of serotinous cones, the researchers concluded that, “in the presence of pine squirrels, the frequency of serotiny is lower and more variable, presumably reflecting,” among a variety of other factors, “the strength of selection exerted by pine squirrels.”
A study published in PNAS in 2014 added evidence to this conclusion. While acknowledging that fire plays a major role in the frequency of serotinous cones, the researchers asserted that “squirrels select against serotiny and that the strength of selection increases with increasing squirrel density.” However, despite making it easier for squirrels to access their seeds, lodgepole pines maintain a degree of serotinous cones, since clearly their main advantage is retaining a canopy-level seed bank from which seeds are released after a fire and by which a new generation of lodgepole pines is born.
Gardening was my first introduction to plants. I enjoyed growing plants so much that I decided to study them. Or rather, I studied the growing of them, i.e. horticulture. During my studies, I became increasingly interested in botany, a vast scientific field that investigates all things plant related, from their evolutionary history to their cellular biology to their interactions with other organisms, etc. Now I am obsessed with pretty much anything to do with botany. However, the molecular side of plant science has never been much of a pursuit of mine. Until now.
What has piqued my interest in this isn’t a university course or a dense textbook on the subject, but instead a podcast hosted by two molecular biologists – Tegan and Joram – who make learning about molecular plant science considerably more interesting than I had previously found it to be. Their podcast (and blog of the same name) is called Plants and Pipettes, and they have been consistently publishing both written and audio content on their site for well over a year now.
The bulk of the Plants and Pipettes podcast consists of Tegan and Joram summing up and discussing a recent plant science research article. While I occasionally get lost in the discussion (particularly when the research delves deep into molecular biology), they both do an exceptional job explaining the science and offering insights that I would not get if I attempted to read the papers on my own. When listening to this portion of the podcast, it helps to have a basic understanding of molecular biology, but it isn’t entirely necessary as the hosts often review basic concepts while discussing the research.
Over the course of the podcast’s history, additional segments have been added. These rarely have anything to do with molecular biology, so if you don’t see yourself tuning in for the research discussion, definitely tune in for the rest. One segment is called My Favorite Plant in which one of the hosts talks about a plant they are interested in that week. Next is Diversity in Plant Science, in which they pick a person that is not a white male and talk about their life and contributions to science (George Washington Carver, for example). After that they define and discuss a cognitive bias, and then they share random things (sometimes science-y, sometimes not) that they find fun or interesting or important to share. Each episode typically ends with a cat fact, as they both have a profound love for cats (although everything is a cat to Joram, apparently).
grass triggerplant (Stylidium graminifolium) was Joram’s favorite plant in episode 12 of Plants and Pipettes (image credit: wikimedia commons)
A highlight among the early episodes was an interview they did with a researcher at the University of Minnesota who is working with pennycress (Thlaspi arvense). This plant is a common weed, but it shows potential for being a productive and useful oilseed crop, similar to a few of its relatives in the mustard family. Speaking of weeds, a fun fact in episode 29 caught my interest, in which Tegan shares an example of Vavilovian mimicry involving rice and barnyard grass (Echinochloa crus-galli). A great introduction to their ongoing series about cognitive biases is episode 37 in which they discuss the Texas sharpshooter fallacy. And of course, I have to recommend listening to episode 48, in which Tegan gives a shout out to Awkward Botany and my new zine Dispersal Stories. How cool is that!?
pennycress (Thlaspi arvense) discussed in episode 8 of Plants and Pipettes (image credit: wikimedia commons)
While I am not always able to keep up with the discussions about molecular plant biology, I still really love listening to this podcast. Apart from the interesting content, the hosts are the real appeal. Not only do I appreciate their social justice rants and their support for open science, but I also find their sense of humor and lack of pretension refreshing. They are excellent models of the way that science communication should be done.
If you check out Plants and Pipettes and decide you need more Tegan and Joram in your life, check out a new podcast they just started with Ellen from Plant Crimes podcast called Plant Book Club, in which they choose a plant-themed book to read and discuss. You can also watch/listen to Tegan and Joram talking about their podcast on Career Conversations.
The main way that a plant gets from place to place is in the form of a seed. As seeds, plants have the ability to travel miles from home, especially with the assistance of outside forces like wind, water, and animals. They could also simply drop to the ground at the base of their parent plant and stay there. The possibilities are endless, really.
But what about plants that don’t even bother making seeds? How do they get around? In the case of bulbous bluegrass, miniature bulbs produced in place of flowers function exactly like seeds. They are formed in the same location as seeds, reach maturity and drop from the plant just like seed-bearing fruits, and are then dispersed in the same ways that seeds are. They even experience a period of dormancy similar to seeds, in that they lie in wait for months or years until the right environmental conditions “tell” them to sprout. And so, bulbils are basically seeds, but different.
bulbous bluegrass (Poa bulbosa)
Bulbous bluegrass (Poa bulbosa) is a Eurasian native but is widely distributed outside of its native range having been repeatedly spread around by humans both intentionally and accidentally. It’s a short-lived, perennial grass that can reach up to 2 feet tall but is often considerably shorter. Its leaves are similar to other bluegrasses – narrow, flat or slightly rolled, with boat-shaped tips and membranous ligules– yet the plants are easy to distinguish thanks to their bulbous bases and the bulbils that form in their flower heads. Their bulbous bases are actually true bulbs, and bulbous bluegrass is said to be the only grass species that has this trait. Just like other bulb-producing plants, the production of these basal bulbs is one way that bulbous bluegrass propagates itself.
basal bulbs of bulbous bluegrass
Bulbous bluegrass is also propagated by seeds and bulbils. Seeds form, like any other plant species, in the ovary of a pollinated flower. But sometimes bulbous bluegrass doesn’t make flowers, and instead modifies its flower parts to form bulbils in their place. Bulbils are essentially tiny, immature plants that, once separated from their parent plant, can form roots and grow into a full size plant. The drawback is that, unlike with most seeds, no sexual recombination has occurred, and so bulbils are essentially clones of a single parent.
The bulbils of bulbous bluegrass sit atop the glumes (bracts) of a spikelet, which would otherwise consist of multiple florets. They have dark purple bases and long, slender, grass-like tips. Bulbils are a type of pseudovivipary, in that they are little plantlets attached to a parent plant. True vivipary occurs when a seed germinates inside of a fruit while still attached to its parent.
Like seeds, bulbils are small packets of starch and fat, and so they are sought ought by small mammals and birds as a source of food. Ants and small rodents are said to collect and cache the bulbils, which is one way they get dispersed. Otherwise, the bulbils rely mostly on wind to get around. They then lie dormant for as long as 2 or 3 years, awaiting the ideal time to take root.
bulbils of bulbous bluegrass
Bulbous bluegrass was accidentally brought to North America as a contaminant in alfalfa and clover seed. It was also intentionally planted as early as 1907 and has been evaluated repeatedly by the USDA and other organizations for use as a forage crop or turfgrass. It has been used in restoration to stabilize soils and reduce erosion. Despite numerous trials, it has consistently underperformed mainly due to its short growth cycle and long dormancy period. It is one of the first grasses to green up in the spring, but by the start of summer it has often gone completely dormant, limiting its value as forage and making for a pretty pathetic turfgrass. Otherwise, it’s pretty good at propagating itself and persisting in locations where it hasn’t been invited and is now mostly considered a weed – a noxious one at that according to some states. Due to its preference for dry climates, it is found most commonly in western North America.
In its native range, bulbous bluegrass frequently reproduces sexually. In North America, however, sexual reproduction is rare, and bulbils are the most common method of reproduction. Prolific asexual reproduction suggests that bulbous bluegress populations in North America should have low genetic diversity. Researchers set out to examine this by comparing populations found in Washington, Oregon, and Idaho. Their results, published in Northwest Science (1997), showed a surprising amount of genetic variation within and among populations. They concluded that multiple introductions, some sexual reproduction, and the autopolyploidy nature of the species help explain this high level of diversity.
———————
Interested in learning more about how plants get around? Check out the first issue of our new zine Dispersal Stories.
Crop domestication often involves selection for larger fruits. In some crops, humans took plant species with relatively small fruits and, over many generations of artificial selection, developed a plant with much larger fruits. Consider giant pumpkins as an extreme example. Yet in the case of apples, relatively large fruits already existed in the wild. Producing larger apples happened quickly and, perhaps even, unconsciously. Apples were practically primed for domestication, and as Robert Spengler explains in a paper published last year in Frontiers in Plant Science, looking back in time at the origins of the apple genus, Malus, can help us understand how the apple we know and love today came to be.
Apples are members of the rose family (Rosaceae), a plant family that today consists of nearly 5000 species. According to the fossil record, plants in the rose family were found in large numbers across North America as early as the Eocene (56 – 33.9 million years ago). They were present in Eurasia at this time as well, but Spengler notes, “there is a much clearer fossil record for Rosaceae fruits and seeds in Europe and Asia during the Miocene and Pliocene (20 – 2.6 million years ago).” Around 14 million years ago, larger fruits and tree-form growth habits evolved in Rosaceae subfamilies, giving rise to the genera Malus and Pyrus (apples and pears). Small, Rosaceae fruits were typically dispersed by birds, but as Sprengler writes, “it seems likely that the large fruits [in Malus and Pyrus] were a response to faunal dispersers of the late Miocene through the Pliocene of Eurasia.” Larger animals were being recruited for seed dispersal in a changing landscape.
Glacial advances and retreats during the Pleistocene (2.6 million – 11,700 years ago) brought even more changes. Plants with effective, long distance seed dispersal were favored because they were able to move into glacial refugium during glacial advances. Even today, these glacial refugium are considered genetic hot spots for Malus, and could be useful for future apple breeding. As the Pleistocene came to a close, many megafauna were going extinct. This continued into the Holocene. Large-fruited apple species lost their primary seed dispersers, and their ranges became even more contracted.
Humans have had an extensive relationship with apples, which began long before domestication. Foraging for apples was common, and seeds were certainly spread that way (perhaps even intentionally). Favorable growing conditions were also created when forests were cleared and old fields were left fallow. Apple trees are early successional species that easily colonize open landscapes, gaps in forests, and forest edges, so human activity that would have created such conditions “could have greatly promoted the spread and success of wild Malus spp. trees during the Holocene.”
The earliest evidence we have of apple domestication (in which “people were intentionally breeding and directing reproduction”) occurred around 3000 years ago in the Tian Shan Mountains of Kazakhstan, where Malus sieversii – a species that is now facing extinction – was being cultivated. This species was later brought into contact with other apple species, a few of which were also being cultivated, including M. orientalis, M. sylvestris, and M. baccata. These species easily hybridized, giving us the modern, domesticated apple, M. domestica. As Spengler writes, “the driving force of apple domestication appears to have been the trans-Eurasian crop exchange, or the movement of plants along the Silk Road.” Continued cultivation and further hybridization among M. domestica cultivars over the past 2000 years has resulted in thousands of different apple varieties.
The unique thing about domesticated apples is that their traits are not fixed in the same way that traits of other domesticated crops are. Growing an apple from seed will result in a very different apple than the apple from which the seed came. Apple traits instead have to be maintained through cloning, which is accomplished mainly through cuttings and grafting. Apples hybridize with other apple species so readily that most apple trees found in the wild are hybrids between wild and cultivated populations.
Spengler considers the study of apple domestication to be “an important critique of plant domestication studies broadly, illustrating that there is not a one-size fits-all model for plant domestication.” The “key” for understanding apple domestication “rests in figuring out the evolutionary driver for large fruits in the wild – seed dispersal through megafaunal mammals – and the process of evolution for these large fruits – hybridization.” He notes that “domestication studies often ignore evolutionary processes leading up to human cultivation,” which, in the case of apples, involves “hybridization events in the wild” that led to the evolution of large fruits “selected for through the success in recruiting large megafaunal mammals as seed disperses.” Many of those mammals went extinct, but humans eventually assumed the role, selecting and propagating “large-fruiting hybrids through cloning and grafting – creating our modern apple.”
Indeed, the relationship between apples and people is close and complex, spanning at least five millennia. The story of the apple begins along the Silk Road… In recent years genetic studies have resolved much of the debate over these origins. Nevertheless, the ancestry of the apple is highly complex. Cloning, inbreeding, and reproduction between species have created a genealogy that looks more like a spider’s web than a family tree. To growers, the beauty of the apple lies not in its rosy skin but in its genetic variability and plasticity, its ability to cross with other species of Malus and other distant lines of M. domestica, and the ease with which it can be grafted onto different rootstocks and cloned.