Pollination in flowering plants is the process of moving pollen grains, which carry sperm cells, from the anthers to the stigmas of either the same flower or a separate flower. If things go well from there, sperm cells will be transported via pollen tubes into the ovaries where fertilization with egg cells can take place and seeds can form. Pollen grain development occurs within the anthers, and by the time the anthers dehisce – or split open – they are ready for transport.
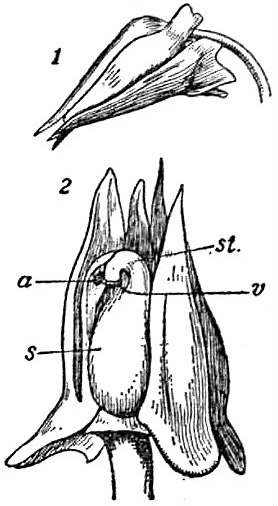
In order to protect the enclosed sperm cells and aid in their movement, pollen grains consist of a series of layers that, among other things, help ensure safe travel. Two major layers are an internal layer called intine, composed largely of cellulose, and an external layer called exine, composed mainly of sporopollenin (a highly durable and complex biopolymer). In many flowering plants, especially those that rely on animals to help carry their pollen, an additional outer layer called pollenkitt is added to the pollen grains before anthers dehisce.
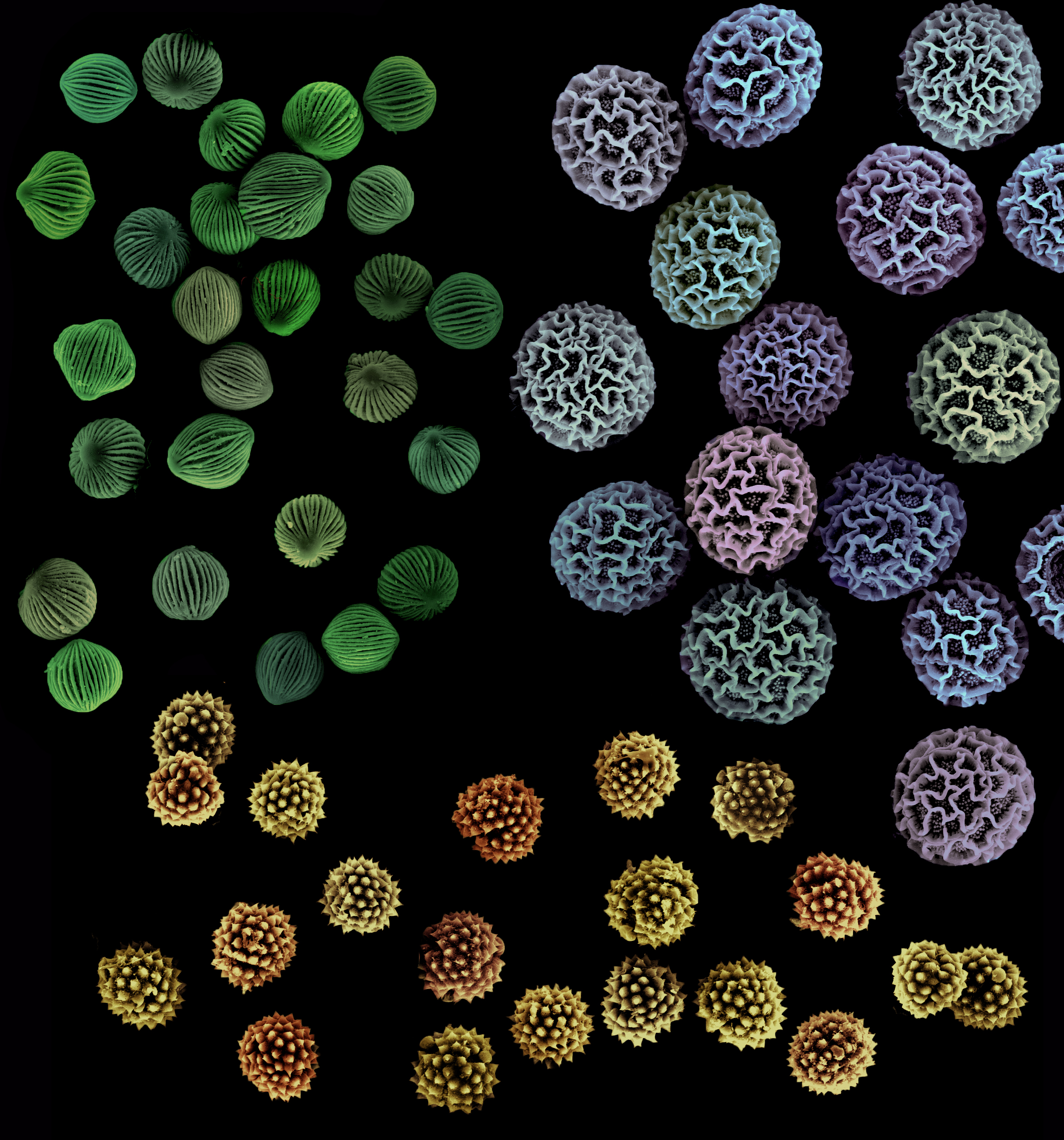
Pollenkitt is an oily, viscous, hydrophobic layer composed of lipids, carotenoids, flavonoids, proteins, and carbohydrates derived from the breakdown of an internal layer of the anther called the tapetum. Pollenkitt forms a sticky layer around the pollen grains and can add color to the pollen other than the typical yellow. The thickness of the pollenkitt and its composition is species specific. In fact, the look, size, and shape of pollen grains themselves are unique to each species and can even be used to help identify plants. Pollenkitt is found in almost all families of flowering plants and is particularly prevalent in species that are animal-pollinated. One exception is the mustard family (Brassicaceae), whose pollen grains are coated in a substance known as tryphine, which functions similar to pollenkitt but whose formation and composition differ enough to be considered separately.

The sticky nature of pollenkitt has numerous functions. For one, it helps pollen grains remain on anthers until an animal comes along to remove them. It also holds pollen grains together in clumps, helps pollen grains stick to insect (and other animal) pollinators during transport, and helps adhere them to stigmas when deposited. A paper published in Flora (2005) lists twenty possible functions for pollenkitt, many of which have been confirmed in certain species and some of which are hypothetical. In addition to functions having to do with pollen movement and placement, pollenkitt may also provide protection from water loss, UV radiation, and fungal and bacterial invasions. In species where pollen is offered as food to pollinating insects, pollenkitt is a more easily digestible food source than the pollen grain itself. Thanks to carotenoids, pollenkitt can make pollen more colorful, which may help attract pollinating insects, or, depending on the color, can also hide pollen from insect visitors.
Another important function of pollenkitt is to give pollen a scent. Odors can help encourage insect visitors or deter them, so depending on the situation, scented pollenkitt may be attracting pollinators or discouraging pollen consumers. In a study published in American Journal of Botany (1988), Heidi Dobson analyzed the chemical composition of 69 different species of flowering plants. She isolated numerous scent compounds in pollenkitt and suggested that “some of the chemicals in pollenkitt may … serve as identification cues to pollen-foraging bees.” Most of the species she analyzed were pollinated by bees (which consume pollen), but the few that were mainly pollinated by hummingbirds and butterflies tended to have fewer scent compounds. Since birds and butterflies are there for the nectar and not the pollen, it would make sense that the pollen of these plant species wouldn’t need to carry a scent.

In flowers that are wind-pollinated, the pollenkitt layer is either very thin or absent altogether. In this case, pollen grains need to be easily released from the anther and are better off when they aren’t sticking to other pollen grains. That way, they are free to be carried off in the breeze to nearby flowers. Some plant species are amphiphilous, meaning they can be both animal-pollinated and wind-pollinated, and according to the authors of the paper published in Flora (2005), pollenkitt layers in these species exhibit intermediate characteristics of both types of pollen grains, generally with thinner, less-sticky pollenkitt and more pollenkitt found within the cavities of the exine.
It’s clear that this unique pollen-glueing substance plays a critical role in the pollination process for many plant species. Considering that each species of plant has its own story to tell, there is still more to learn about the forms and functions that pollenkitt takes.
———–
This is the first in a series of posts in 2024 in which, once again, I am exploring the world of pollinators and pollination. You can read more about this effort in last month’s Year in Review post.