The lettuce we eat is a close relative to the lettuce we weed out of our gardens. Last week we discussed the potential that wild relatives may have for improving cultivated lettuce. But if wild lettuce can be crossed with cultivated lettuce to create new cultivars, can cultivated lettuce cross with wild lettuce to make it more weedy?
Because so many of our crops are closely related to some of the weeds found along with them or the plants growing in nearby natural areas, the creation of crop-wild hybrids has long been a concern. This concern is heightened in the age of transgenic crops (also known as GMOs), for fear that hybrids between weeds and such crops could create super weeds – fast spreading or highly adapted weeds resistant to traditional control methods such as certain herbicides. To reduce this risk, extensive research is necessary before such crops are released for commercial use.

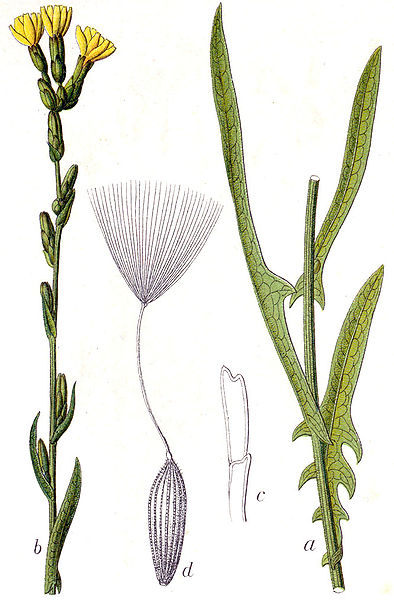
flowers of prickly lettuce (Lactuca serriola)
There are no commercially available, genetically modified varieties of cultivated lettuce, so this is not a concern when it comes to crop-wild hybrids; however, due to how prevalent weedy species like prickly lettuce (Lactuca serriola) are, hybridization with cultivated lettuce is still a concern. So, it is important to understand what the consequences might be when hybridization occurs.
In a paper published in Journal of Applied Ecology in 2005, Hooftman et al. examined a group of second-generation hybrids (L. sativa x L. serriola), and found that the hybrids behaved and appeared very similarly to non-hybrid prickly lettuce. They also found that the seeds produced by the hybrids had a significantly higher germination rate than non-hybrid plants. This is an example of hybrid vigor. Thus, “if hybridization does occur, this could lead to better performing and thus potentially more invasive (hybrid) genotypes.” However, the authors cautioned that “better performing genotypes do not automatically result in higher invasiveness,” and that much depends on the conditions they are found in, the level of human disturbance, etc.
Another thing to consider is that hybrids are not stable. In an article published in Nature Reviews Genetics in 2003, Stewart et al. adress the “misunderstanding that can arise through the confusion of hybridization and … introgression.” It is wrong to assume that hybrids between crops and wild relatives will automatically lead to super weeds. For this to occur, repeated crosses with parental lines (also known as backcrossing) must occur, and “backcross generations to the wild relative must progress to the point at which the transgene [or other gene(s) in question] is incorporated into the genome of the wild relative.” That is what is meant by “introgression.” This may happen quickly or over many generations or it may never happen at all. Each case is different.

prickly leaf of prickly lettuce (Lactuca serriola)
In a paper published in Journal of Applied Ecology in 2007, Hooftman et al. observe the breakdown of crop-wild lettuce hybrids. They note that “fitness surplus through [hybrid vigor] will often be reduced over few generations,” which is what was seen in the hybrids they observed. One possible reason why this occurs is that lettuce is predominantly a self-crossing species; outcrossing is rare, occurring 1 – 5% of the time thanks to pollinating insects. But that doesn’t mean that a stable, aggressive genotype could never develop. Again, much depends on environmental conditions, as well as rates of outcrossing and other factors relating to population dynamics.
A significant expansion of prickly lettuce across parts of Europe led some to hypothesize that crop-wild hybrids were partly to blame. In a paper published in Molecular Ecology in 2012 Uwimana et al. ran population genetic analyses on extensive data sets to determine the role that hybridization had in the expansion. They concluded that, at a level of only 7% in wild habitats, crop-wild hybrids were not having a significant impact. They observed greater fitness in the hybrids, as has been observed in other studies (including the one above), but they acknowledged the instability of hybrids, especially in self-pollinating annuals like lettuce.

seed head of prickly lettuce (Lactuca serriola)
It is more likely that the expansion of prickly lettuce in Europe is due to “the expansion of favorable habitat as a result of climate warming and anthropogenic habitat disturbance and to seed dispersal because of transportation of goods.” Uwimana et al. did warn, however, that “the occurrence of 7% crop-wild hybrids among natural L. serriola populations is relatively high [for a predominantly self-pollinating species] and reveals a potential [for] transgene movement from crop to wild relatives [in] self-pollinating crops.”