Desert conditions are not intuitively conducive to life. In many regards they are extreme. Blistering, bleak, dry, and barren. The desert is a place unsuited for the faint of heart and the ill-equipped. Broadly speaking, life in the desert is reliant on one of two things: technology or evolutionary adaptation. Like many species native to desert environments, the desert tortoise employs the latter. It is at home in the desert because it evolved there. That is not to say that life is always easy for the desert tortoise and species like it, but it is possible, thanks to hundreds of thousands of years of making it work. As John Alcock puts it in, Sonoran Desert Spring, “the tortoise will deal with its environment through evolved design rather than seek to deny the desert its due.”
Perhaps it is because the desert is such a harsh environment, requiring finely tuned adaptations for survival, that sweeping changes can put resident species in peril – threatening their long-term existence. The desert tortoise is an example of this. Since 1990, Gopherus agassizii has found itself listed as threatened under the U.S. Endangered Species Act [it is categorized as vulnerable by the IUCN] due to significant population declines and loss of habitat. Getting there did not happen overnight, and it is impossible to pinpoint a sole cause of the tortoise’s decline. Instead, a suite of things have conspired against it, making it difficult to decide on the best route towards conservation.
In an October 2012 issue of BioScience, Averill-Murray, et al. enumerate some of the human-medaited threats that act both simultaneously and synergistically against desert tortoise populations:
Habitat conversion occurs as a result of urban development, mining, waste disposal, energy development, and road construction. Habitat modification is caused by military training, off-highway vehicle use, utility corridors, livestock grazing, and the proliferation of invasive plants. … Direct losses of tortoises also occur through predation, disease, collection from the wild, and recreational killing.
Apart from climate change, which is projected to substantially reduce the historical range of the desert tortoise in the coming years, the proliferation of introduced grasses is particularly disconcerting. Such grasses tend to increase wildfire frequency in areas where wildfire is historically rare and the native flora is ill-adapted to frequent fire. This can alter plant communities in a way that favors introduced plants over plants native to the region.
The desert tortoise is the largest terrestrial turtle in the United States, measuring up to 15 inches long and weighing up to 15 pounds. Their carapaces are generally dull brown or gray, although those of young tortoises may have orange markings. Their limbs are stocky and elephantine, and their front legs are shovel-like and equipped with claws for digging. They reach sexual maturity between ages 15-20, generally living for at least 35 years and as many as 50-100 years.
Desert tortoises are distributed throughout the Mohave and Sonoran Deserts of southeastern United States and into the Sonaron Desert and Sinaloan foothills of northwestern Mexico. Their habitat varies widely across their range. In general, tortoises prefer sites where the soil is loamy and easy to dig as they spend much of their time in underground dens; however, they also occur in rocky foothills where shelter can be found among the rocks. In the Mohave Desert, they are commonly found in plant communities that are dominated by creosote bush (Larrea tridentata), which they use for shade and an occasional food source.
Recently the species known as Gopherus agassizii was determined to consist of at least two (possibly four) distinct species. Desert tortoises that occur north and west of the Colorado River have retained the scientific name G. agassizii and are commonly referred to as Agassiz’s desert tortoise. Desert tortoises occurring east of the Colorado river have been given the name G. morafkai, commonly known as Morafka’s desert tortoise. In light of this, G. agassizii may find itself uplisted to endangered, as its range has been reduced to about 30% of its former self and its southern cousins can no longer be considered a genetic reservoir.
Seeing that desert tortoises have plenty of the right foods to eat ensures their immediate survival and holds them back from the precipice of extinction. The question, “What does a desert tortoise eat?,” was what peaked my curiosity in this subject to begin with. I knew they were herbivores (for the most part), so I assumed they must have a favorite food – something that composed the majority of their diet. Finding an answer to this question led me down a rabbit hole [or should I say a tortoise hole? Some tortoise dens can extend 30 feet or more into the banks of desert dry washes.] that led me to discover the complexity of these creatures. It turns out, there is no easy answer to my initial question. What a desert tortoise eats depends on where in its expansive range it resides, what time of year it is, what plants are available in a particular year, whether or not it’s a drought year, etc.
The desert tortoise is “one of the most studied reptiles in the world,” so hundreds of observations have been made, leading to dozens of reports and studies that examine the diet of the desert tortoise; however, the results are highly variable. Due to such variability, this fact sheet from the San Diego Zoo states matter-of-factly, “an ‘average’ tortoise diet [is] hard to characterize.” But let’s try.
The desert tortoise emerges from its winter den in early spring. At the same time, annual wildflowers are also emerging, taking advantage of warming temperatures and rare soil moisture accumulated during winter precipitation. This is the desert tortoise’s preferred banquet. Because there will be little water available the rest of the year, desert tortoises hydrate themselves mainly through the plants they eat. The lush stems, leaves, and flowers of annual wildflowers provide both nutrients and the water necessary to sustain themselves throughout much of the year and aid in their growth and reproduction.
As spring turns to summer, the tortoises switch to eating herbaceous perennials and grasses. By this point, both introduced annual grasses and native perennial bunchgrasses are drying up, but tortoises are still able to extract some nutrients and moisture by eating their dry stems and leaves. Cactus pads and fruits (particularly those in the genus Opuntia) as well as young leaves of shrubs also help tortoises subsist through the long, hot summers, which are mostly spent deep in their dens away from predators and the blistering heat.
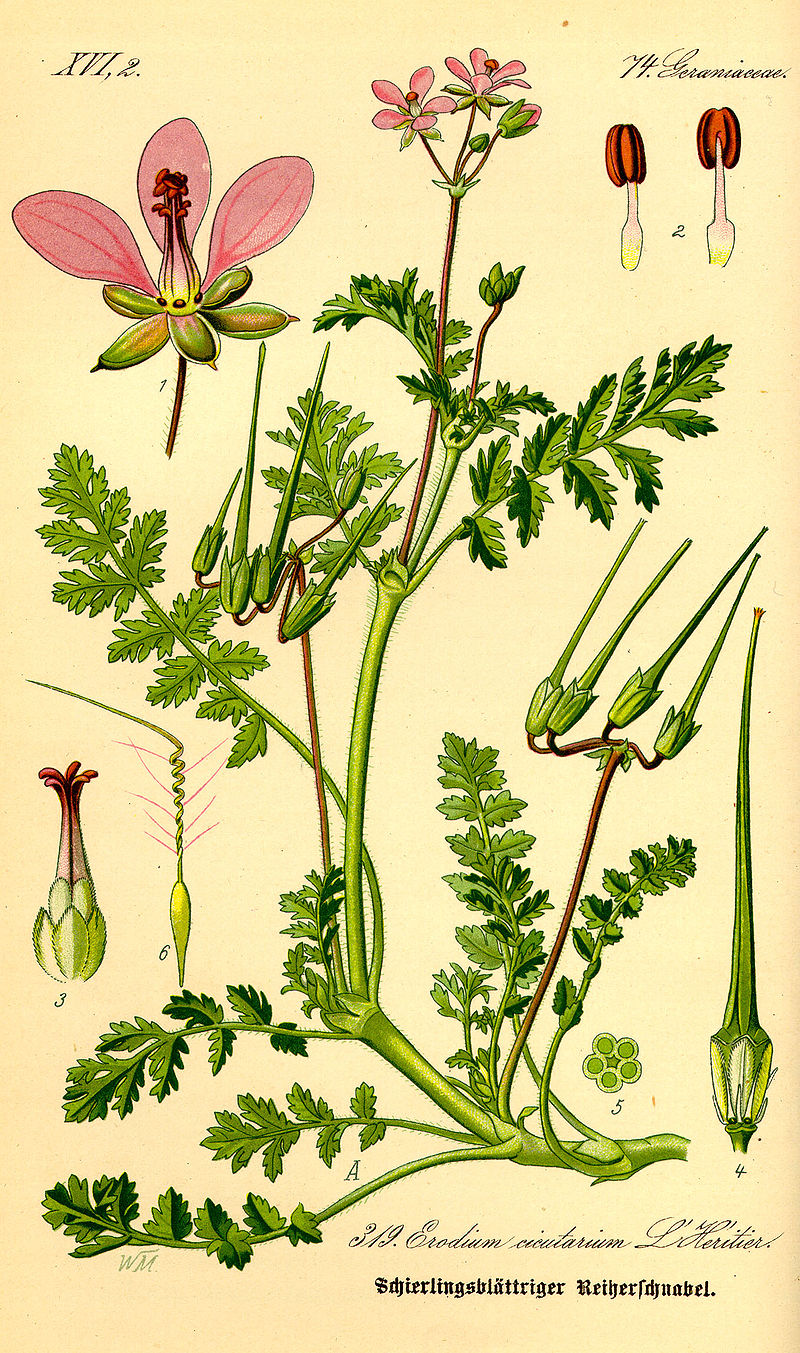
A paper published in a March 1986 issue of Herpetologica follows a group of tortoises over the period of a year and makes a number of lifestyle observations, including their diet. The authors noted that much of their diet consisted of two annual wildflowers (Camissonia munzii and Langloisia setosissima), a perennial bunchgrass (Achnatherum hymenoides), and a non-native annual grass (Bromus rubens). A paper published in a 2010 issue of Journal of Herpetology compared the nutritional quality of four plant species commonly consumed by desert tortoises: a native and non-native grass (Achnatherum hymenoides and Schismus barbatus) and a native and non-native annual forb (Malacothrix glabrata and Erodium cicutarium). They found little difference between the native and non-native species in either catagory, but determined that the forbs were significantly more nutritious than the grasses, which lead them to recommend managament practices that would increase the availability of forbs (regardless of provenance) in tortoise habitat. Numerous studies have documented the frequent consumption of introduced plant species by desert tortoises.

Redstem stork’s bill (Erodium cicutarium) is an introduced species commonly consumed by desert tortoises – image credit: wikimedia commons
For me, one of the most interesting things to learn was the variety of “non-food” items that tortoises may consume. Tortoises are often observed eating soil and rocks, and are also known to eat bones, arthropods, feces, feathers, hair, and egg shells. The rocks are thought to act as a gastrolith, aiding in digestion. The other items may help supplement minerals and nutrients the tortoises are lacking in their plant-based diet, particularly calcium which is greatly needed for growth and reproduction. Shockingly, a report that appeared in a 2007 issue of The Southern Naturalist details incidences of tortoises eating the skeletal remains of other tortoises.
Desert tortoises are an engrossing subject of study, and so much more could be said about them. For now, I leave you with this passage from Alcock’s book:
To see a tortoise with wrinkled neck and solemn eyes, moving like an animated rock, is an essential part of the experience of the desert. The removal of even a single adult extinguishes a presence that was meant to persist for years to come and snuffs out a prehistoric spark of life in a spartan environment where life, so hard-won, should be celebrated.
Additional Resources: