
What Is a Plant?
Part one and two of this series have hopefully answered that.
Why should you care?
Part three offered a pretty convincing answer: “if it wasn’t for [plants], there wouldn’t be much life on this planet to speak of.”
Plants are at the bottom of the food chain and are a principle component of most habitats. They play major roles in nutrient cycling, soil formation, the water cycle, air and water quality, and climate and weather patterns. The examples used in part three of this series to explain the diverse ways that plants provide habitat and food for other organisms apply to humans as well. However, humans have found numerous other uses for plants that are mostly unique to our species – some of which will be discussed here.
But first, some additional thoughts on photosynthesis. Plants photosynthesize thanks to the work accomplished by very early photoautotrophic bacteria that were confined to aquatic environments. These bacteria developed the metabolic processes and cellular components that were later co-opted (via symbiogensis) by early plants. Plants later colonized land, bringing with them the phenomena of photosynthesis and transforming life on earth as we know it. Single-celled organisms started this whole thing, and they continue to rule. That’s just something to keep in mind, since our focus tends to be on large, multi-cellular beings, overlooking all the tiny, less visible beings at work all around us making life possible.
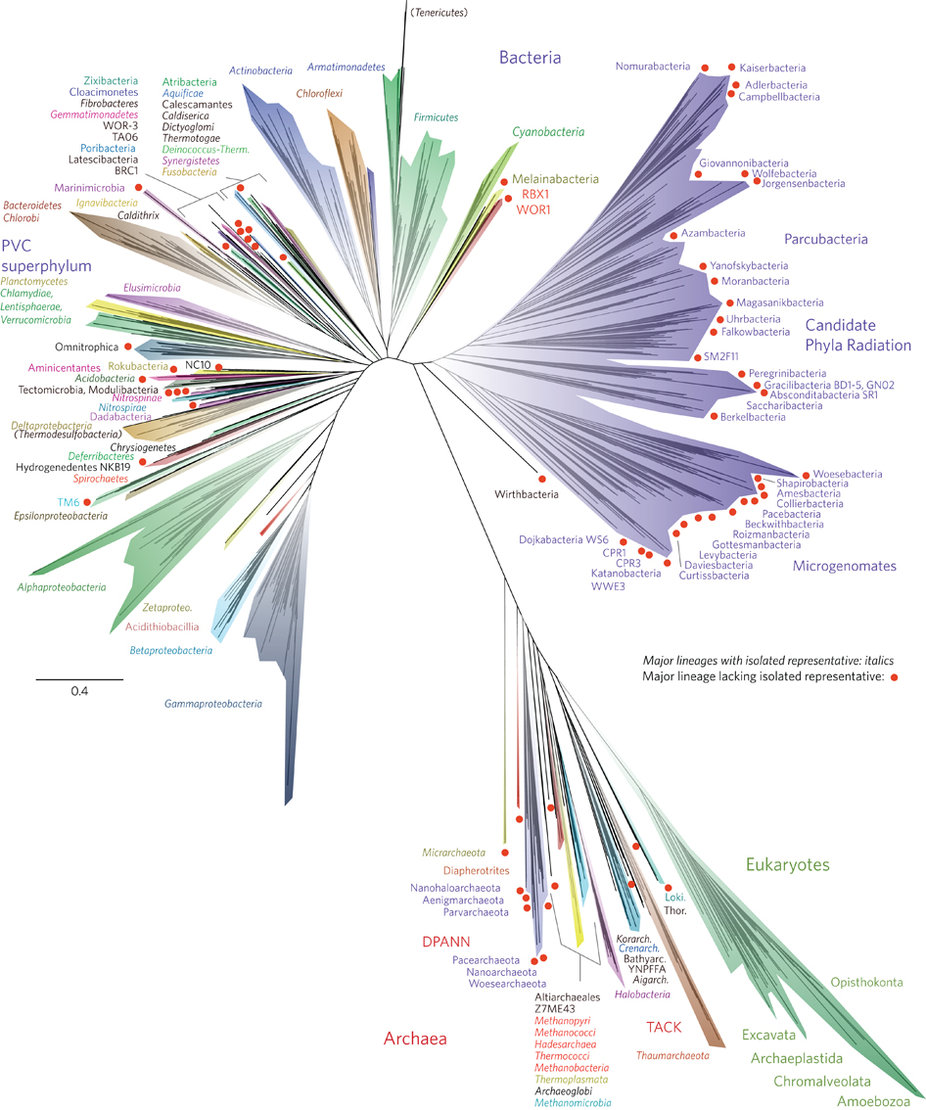
Current representation of the tree of life. Microorganisms clearly dominate. (image credit: nature microbiology)
Food is likely the first thing that comes to mind when considering what use plants are to humans. The domestication of plants and the development of agriculture are easily among the most important events in human history. Agricultural innovations continue today and are necessary in order to both feed a growing population and reduce our environmental impact. This is why efforts to discover and conserve crop wild relatives are so essential.
Plants don’t just feed us though. They house us, clothe us, medicate us, transport us, supply us, teach us, inspire us, and entertain us. Enumerating the untold ways that plants factor in to our daily lives is a monumental task. Rather than tackling that task here, I’ll suggest a few starting points: this Wikipedia page, this BGCI article, this Encylopedia of Life article, and this book by Anna Lewington. Learning about the countless uses humans have found for plants over millennia should inspire admiration for these green organisms. If that admiration leads to conservation, all the better. After all, if the plants go, so do we.

Humans have a long tradition of using plants as medicine. Despite all that we have discovered regarding the medicinal properties of plants, there remains much to be discovered. This is one of the many reasons why plant conservation is imperative. (photo credit: wikimedia commons)
Gaining an appreciation for the things that plants do for us is increasingly important as our species becomes more urban. Our dense populations tend to push plants and other organisms out, yet we still rely on their “services” for survival. Many of the functions that plants serve out in the wild can be beneficial when incorporated into urban environments. Plants improve air quality, reduce noise pollution, mitigate urban heat islands, help manage storm water runoff, create habitat for urban wildlife, act as a windbreak, reduce soil erosion, and help save energy spent on cooling and heating. Taking advantage of these “ecosystem services” can help our cities become more liveable and sustainable. As the environmental, social, and economic benefits of “urban greening” are better understood, groups like San Francisco’s Friends of the Urban Forest are convening to help cities across the world go green.
The importance of plants as food, medicine, fuel, fiber, housing, habitat, and other resources is clear. Less obvious is the importance of plants in our psychological well being. Numerous studies have demonstrated that simply having plants nearby can offer benefits to one’s mental and physical health. Yet, urbanization and advancements in technology have resulted in humans spending more and more time indoors and living largely sedentary lives. Because of this shift, author Richard Louv and others warn about nature deficit disorder, a term not recognized as an actual condition by the medical community but meant to describe our disconnect with the natural world. A recent article in BBC News adds “nature knowledge deficit” to these warnings – collectively our knowledge about nature is slipping away because we don’t spend enough time in it.
The mounting evidence for the benefits of having nature nearby should be enough for us to want to protect it. However, recognizing that we are a part of that nature rather than apart from it should also be emphasized. The process that plants went through over hundreds of millions of years to move from water to land and then to become what they are today is parallel with the process that we went through. At no point in time did we become separate from this process. We are as natural as the plants. We may need them a bit more than they need us, but we are all part of a bigger picture. Perhaps coming to grips with this reality can help us develop greater compassion for ourselves as well as for the living world around us.































