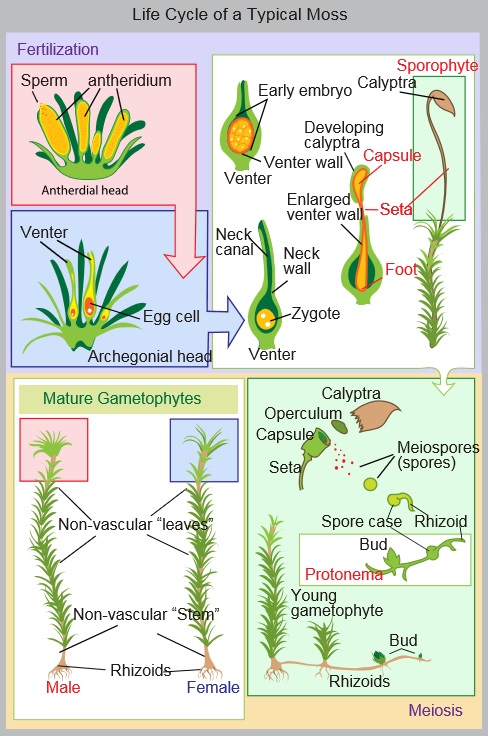
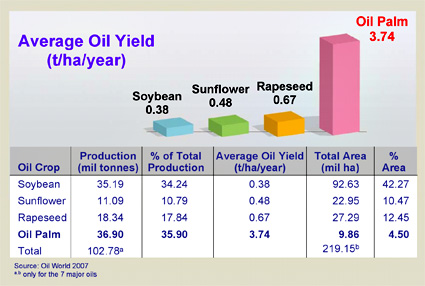
When I decided to spend a year writing about pollinators and pollination, I specifically wanted to focus on pollinators besides the honey bee. Honey bees already get lots of attention, and there are loads of other pollinating organisms that are equally fascinating. But that’s just the thing, honey bees are incredibly fascinating. They have a strict and complex social structure, and they make honey – two things that have led humans to develop a strong relationship with them. We have been managing honey bees and exploiting their services for thousands of years, and we have spread them across the planet, bringing them with us wherever we go. In North America, honey bees are used to pollinate a significant portion of our pollinator-dependent crops, despite the fact that they are not native to this continent. In that sense, they are just another domesticated animal, artificially selected for our benefit.
It’s common knowledge that honey bees (and pollinators in general) have been having a rough time lately. Loss of habitat, urbanization, industrial farming practices, abundant pesticide use, and a variety of pests and diseases have been making life difficult for pollinators. Generally, when the plight of pollinators comes up in the news, reference is made to honey bees (or another charismatic pollinator, the monarch butterfly). News like this encourages people to take action. On the positive side, efforts made to protect honey bees can have the side benefit of protecting native pollinators since many of their needs are the same. On the negative side, evidence suggests that honey bees can compete with native pollinators for limited resources and can pass along pests and diseases. Swords are often double-edged, and there is no silver bullet.
In a recent conversation with a budding beekeeper, I was recommended the documentary, More than Honey. I decided to watch it, write a post about it, and call that the honey bee portion of the Year of Pollination. Part way through the movie, another documentary, Vanishing of the Bees, was recommended to me, and so I decided to watch both. Below are some thoughts about each film.

More than Honey
Written and directed by Swiss documentary filmmaker, Markus Imhoof, this beautifully shot, excellently narrated, meandering documentary thrusts viewers into incredibly intimate encounters with honey bees. Cameras follow bees on their flights and into their hives and get up close and personal footage of their daily lives, including mating flights, waggle dances, pupating larvae, flower pollination, and emerging queens. In some scenes, the high definition shots make already disturbing events even more disturbing, like bees dying after being exposed to chemicals and tiny varroa mites crawling around on the bodies of bees infecting them with diseases – wings wither away and bees become too weak to walk. This movie is worth watching for the impressive cinematography alone.
But bees aren’t the only actors. The human characters are almost as fun to watch. A Swiss beekeeper looks out over stunning views of the Alps where he keeps his bees. He follows a long tradition of beekeeping in his family and is very particular about maintaining a pure breed in his hives, going so far as flicking away the “wrong” bees from flowers on his property and crushing the head off of an unfaithful queen. A commercial beekeeper in the United States trucks thousands of beehives around the country, providing pollination services to a diverse group of farms – one of them being a massive almond grove in California. He has been witness to the loss of hundreds of honey bee colonies and has had to become “comfortable with death on an epic scale” – the grueling corporate world grinds along, and there is no time for mourning losses.
Further into the documentary, a woman in Austria demonstrates how she manipulates a colony into raising not just one queen, but dozens. She has spent years breeding bees, and her queens are prized throughout the world. A man in Arizona captures and raises killer bees – hybrid bees resulting from crosses between African and European honey bees (also known as Africanized honey bees). Despite their highly aggressive nature, he prefers them because they are prolific honey producers and they remain healthy without the use of synthetic pesticides.
Probably the darkest moment in the film is watching workers in China hand pollinate trees in an orchard. Excessive pesticide use has decimated pollinator populations in some regions, leaving humans to do the pollinating and prompting the narrator to reflect on the question, “Who’s better at pollinating, man or bees? Science answers with a definite, ‘not man.'”
Also included in the film is an intriguing discussion about bees as a super-organism with a German neuroscientist who is studying bee brains. The narrator sums it up like this: “Without its colony the individual bee cannot survive. It must subordinate its personal freedom for the good of the colony… Could it be that individual bees are like the organs or cells of a body? Is the super-organism as a whole the actual animal?”

Vanishing of the Bees
Colony collapse disorder is a sometimes veiled yet important theme throughout More than Honey, and it was certainly something that drove the creation of the film. In the case of Vanishing of the Bees, colony collapse disorder is the reason for its existence. Narrated by actor, Ellen Page, and produced in part by a film production company called Hive Mentality Films, this movie came out on the heels of the news that bee colonies were disappearing in record numbers throughout the world. It tells the story of colony collapse disorder from the time that it first appeared in the news – one of the film’s main characters is the beekeeper that purportedly first brought attention to the phenomenon – and into the years that followed as scientists began exploring potential causes.
This film contains lots of important information and much of it seems credible, but it is also the type of documentary that in general makes me wary of documentaries. Its purpose goes beyond just trying to inform and entertain; it’s also trying to get you on board with its cause. I may agree with much of what is being said, but I don’t particularly like having my emotions targeted in an effort to manipulate me to believe a certain way. It’s a good idea not to let documentaries or any other type of media form your opinions for you. Consider the claims, do some of your own research and investigation, and then come to your own conclusion. That’s my advice anyway…even though you didn’t ask for it.
That being said, colony collapse disorder is a serious concern, and so I’ll end by going back to More than Honey and leave you with this quote by its narrator:
The massive death of honey bees is no mystery. What’s killing them is not pesticides, mites, antibiotics, incest, or stress, but a combination of all these factors. They are dying as a result of our civilization’s success, as a result of man, who has turned feral bees into docile, domestic animals – wolves into delicate poodles.