Seeds house and protect infant plants. When released from their parent plant, they commence a journey that, if successful, will bring them to a suitable location where they can take up residence (upon germination) and carry out a life similar to that of their parents. Their seed coats (and often – in the case of angiosperms – the fruits they were born in) help direct them and protect them in this journey. Physical and chemical factors inhibit them from germinating prematurely – a phenomenon known as dormancy. Agents of dispersal and mechanisms of dormancy allow seeds to travel through time and space — promises of new plants yet to be realized.
There is rarely a need for a seed to germinate immediately upon reaching maturity. In many cases, such as in temperate climates or in times of drought or low light, germinating too soon could be detrimental. The most vulnerable time in a plant’s life comes when it is a young seedling. Thus, finding the right time and space to get a good start is imperative.

The fruits (and accompanying seeds) of doubleclaw (Proboscidea parviflora) are well equipped for long distance dispersal. (via wikimedia commons)
In rare instances, dispersal via seeds offers little advantage; instead, dispersal of live seedlings or propagules is preferable. For this select group of plants, vivipary is part of the reproductive strategy. In vivipary, seeds lack dormancy. Rather than waiting to be dispersed before germinating, viviparous seeds germinate inside of fruits that are still attached to their parent plants.
Occasionally, seeds are observed germinating inside tomatoes, citrus, squash, and other fruits; however, these fruits are usually overripe and often detached from the plant. In these instances, what is referred to as “vivipary” is not a genetic predisposition or part of the reproductive strategy. It’s just happenstance – a fun anomaly. The type of vivipary discussed in this post is actually quite rare, occurring in only a handful of species and prevalent in a select number of environments.
There are three main types of vivipary: true vivipary, cryptovivipary, and pseudovivipary. In true vivipary, a seed germinates inside the fruit and pushes through the fruit wall before the fruit is released. In cryptovivipary, a seed germinates inside the fruit but remains inside until after the fruit drops or splits open. Pseudovivipary is the production of bulbils or plantlets in the flower head. It does not involve seeds and is, instead, a form of asexual reproduction that will be discussed in a future post.
True vivipary is commonly seen among plant communities located in shallow, marine habitats in tropical or subtropical regions, such as mangroves or seagrasses. The term mangrove is used generally to describe a community of plants found in coastal areas growing in saline or brackish water. It also refers more specifically to the small trees and shrubs found in such environments. While not all mangrove species are viviparous, many of them are.
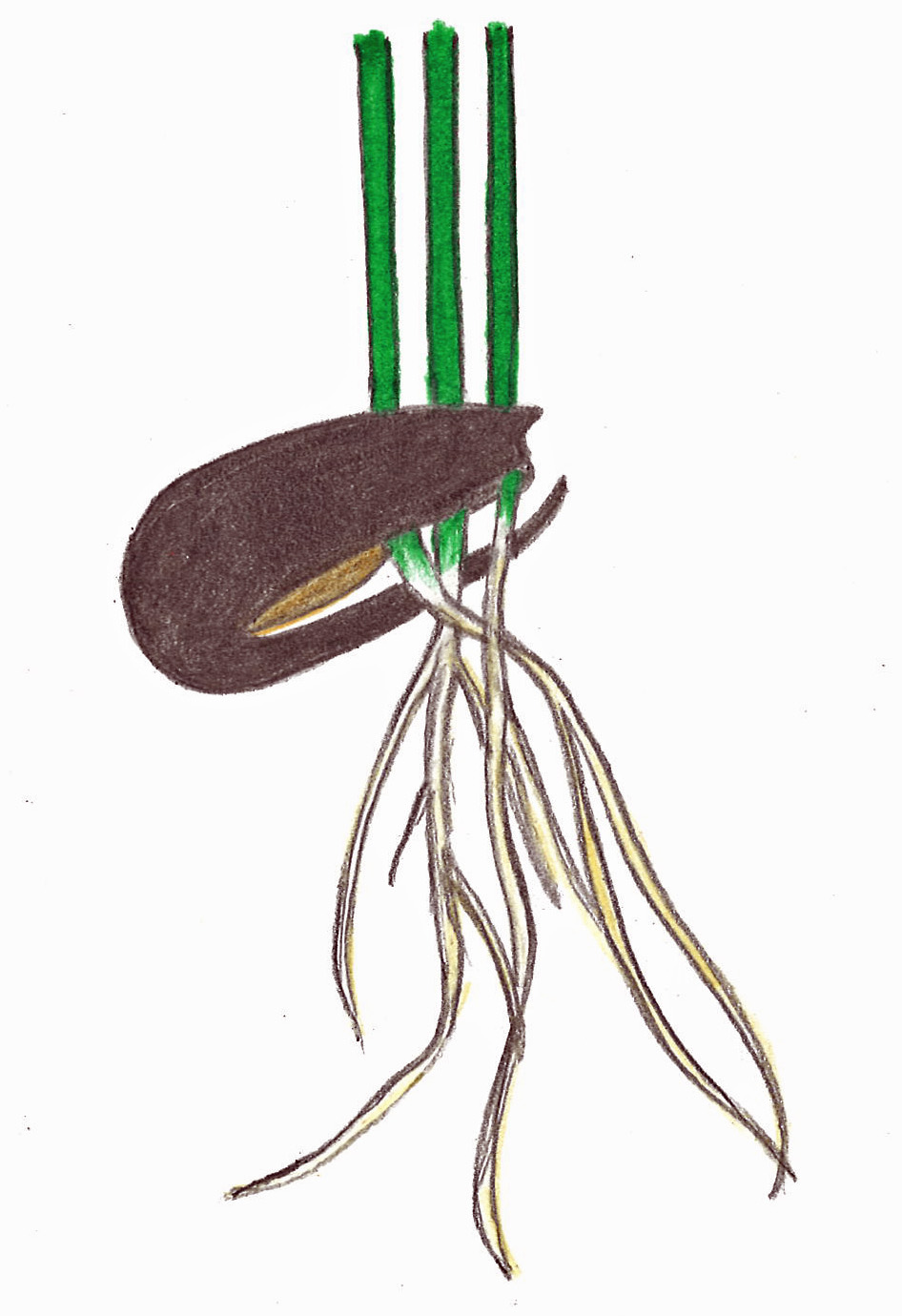
Seedlings of viviparous mangrove species emerge from the fruit and drop from the plant into the salty water below. From there they have the potential to float long or short distances before taking root. They may land in the soil upright, but often, as the tide recedes, they find themselves lying horizontally on the soil. Luckily, they have the remarkable ability to take root and quickly stand themselves up. Doing this allows young plants to keep their “heads” above water as the tides return. It also helps protect the shoot tips from herbivory.

Viviparous seedlings emerging from the fruits of red mangrove (Rhizophora mangle) via wikimedia commons
Another example of vivipary is found in the epiphytic cactus (and close relative of tan hua), Epiphyllum phyllanthus. Commonly known as climbing cactus, this species was studied by researchers in Brazil who harvested fruits at various stages to observe the development of the viviparous seedlings. They then planted the seedlings on three different substrates to evaluate their survival and establishment.
Epiphyllum phyllanthus is cryptoviviparous, so the germinated seeds don’t leave the fruit until after it splits open. In a sense, the mother plant is caring for her offspring before sending them out into the world. The researchers see this as “a form of parental care with subsequent conspecific [belonging to the same species] nursing.” Since the plant is epiphytic – meaning that it grows on the surface of another plant rather than in the soil – local dispersal is important, since there is no guarantee that seeds or propagules dispersed away from the host plant will find another suitable site. That being said, the researchers believe that “vivipary involves adaptation to local dispersal,” since “the greater the dispersal distance is, the higher the risk and the lower the probability of optimal dispersion.”
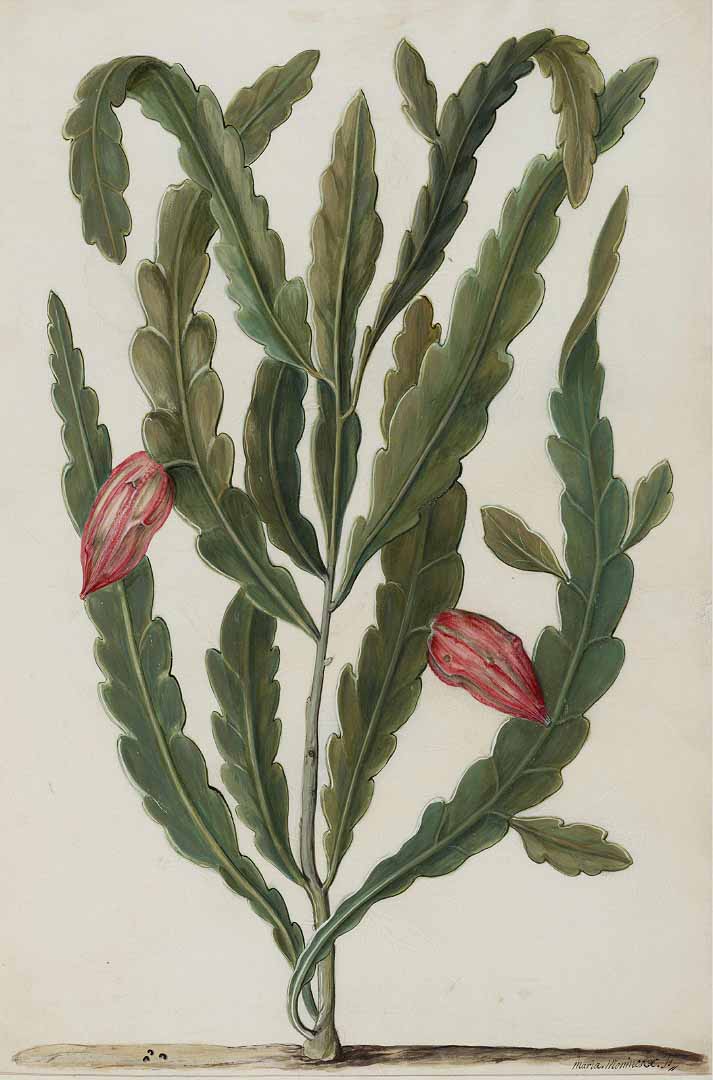
Epiphyllum phyllanthus via Useful Tropical Plants
While some viviparous seedlings of mangroves can travel long distances from their parent plant and don’t always root into the ground immediately, they maintain their advantage over seeds because they can root in quickly upon reaching a suitable site and lift themselves up above rising tide waters. As the authors of the Epiphyllum study put it, vivipary is “a reproductive advantage that, in addition to allowing propagules to root and grow almost immediately, favors quick establishment whenever seedlings land on suitable substrates.”
There is still much to learn about this unusual and rare botanical feature. The research that does exist is relatively scant, so it will be interesting to see what more we can discover. For now, check out the following resources:
- Elmqvist, T. and Cox, P.A.1996. The evolution of vivipary in flowering plants. Oikos. 77(1): 3-9.
- Krauss, K.W. and Ball, M.C. 2013. On the halophytic nature of mangroves. Trees. 27: 7-11.
- Tomlinson, P.B. 2000. Systematic and functional anatomy of seedlings in mangrove Rhizophoraceae: vivipary explained? Botanical Journal of the Linnean Society. 134: 215-231.
- Tomlinson, P.B. 2016. The Botany of Mangroves. Cambridge University Press.
- Guide to the Mangroves of Singapore: Vivipary
Also, check out this You Tube video of :