The theme for the past 15 posts has been the October 2014 Special Issue of American Journal of Botany, Speaking of Food: Connecting Basic and Applied Plant Science. After a brief introduction to the issue, I spent the next 14 posts (spanning a period of 5 weeks) reading and writing summaries of each of the 17 articles. If you actually read every post, you are a champion in my eyes, and I probably owe you a prize of some sort. And even if you just read one or two, thank you, and I hope you found value in what you read.
I have to admit that it was kind of a grueling process. Many of the articles, along with being lengthy, included high level discussions that were beyond my current understanding, especially concerning topics like genetics, genomics, and phylogenetics. I learned a lot while reading them, but I am still far from truly grasping many of the concepts. For that reason, I did not feel completely comfortable writing summaries of some of these discussions. I made an effort not to misrepresent or oversimplify the research, but I can’t say for sure that my attempts were always successful. I welcome any criticisms, corrections, complaints, or comments in this regard, and I am open to making edits or updates to any of the posts as necessary. I consider this blog my learning platform, as well as a place to share my phyto-curiosity. Perhaps you find it a place for learning, too?
The main purpose of this post is to provide a Table of Contents for the last 14 posts, something that will make it easier to navigate through this series without having to scroll through each post. If you are interested in reading the entire series (again, you’re a champion), you can access them all in order here by clicking on the titles. Otherwise, you can pick and choose whatever topics interest you the most.
- On the Origins of Agriculture – A deep dive into plant domestication and the beginnings of agriculture, including the revision of theoretical approaches to thinking about the history of plant domestication and a discussion of emerging methods and tools for exploring early domestication and emerging agriculture.
- The Legacy of a Leaky Dioecy – Does pre-Colombian management of North American persimmon trees explain why non-dioecious individuals are found in an otherwise dioecious species?
- Dethroning Industrial Agriculture: The Rise of Agroecology – The environmentally devastating effects of industrial agriculture can and must be replaced by a more sustainable, ecologically-focused from of agriculture. This will require reforming our economic system and rethinking our “one size fits all” approach to scientific research.
- An Underutilized Crop and the Cousins of a Popular One – Safflower, an underutilized oilseed crop, could be improved by introducing genes from wild relatives. Soybean, a very popular and valuable crop, could also be improved by introducing genes from its perennial cousins.
- Carrots and Strawberries, Genetics and Phylogenetics – An exploration of the genetics and phylogenetics of carrots and strawberries. Better understanding of their genetics will aid in crop improvements; better understanding of their phylogenetics gives us further insight into the evolution of plants.
- Exploring Pollination Biology in Southwestern China – A fascinating look at the pollination biology of edible and medicinal plants in southwestern China, revealing significant gaps in scientific understanding and the need for conservation and continued research.
- Your Food Is a Polyploid – Polyploidy is more prevalent in plants than we once thought. This article examines the role of polyploidy in crop domestication and future crop improvements.
- Tales of Weedy Waterhemp and Weedy Rice – How agriculture influenced the transition to invasiveness in two important weed species.
- Cultivated Sunflowers and Their Wild Relatives – An investigation into the flowering times of wild sunflowers reveals potential for improvements in cultivated sunflowers.
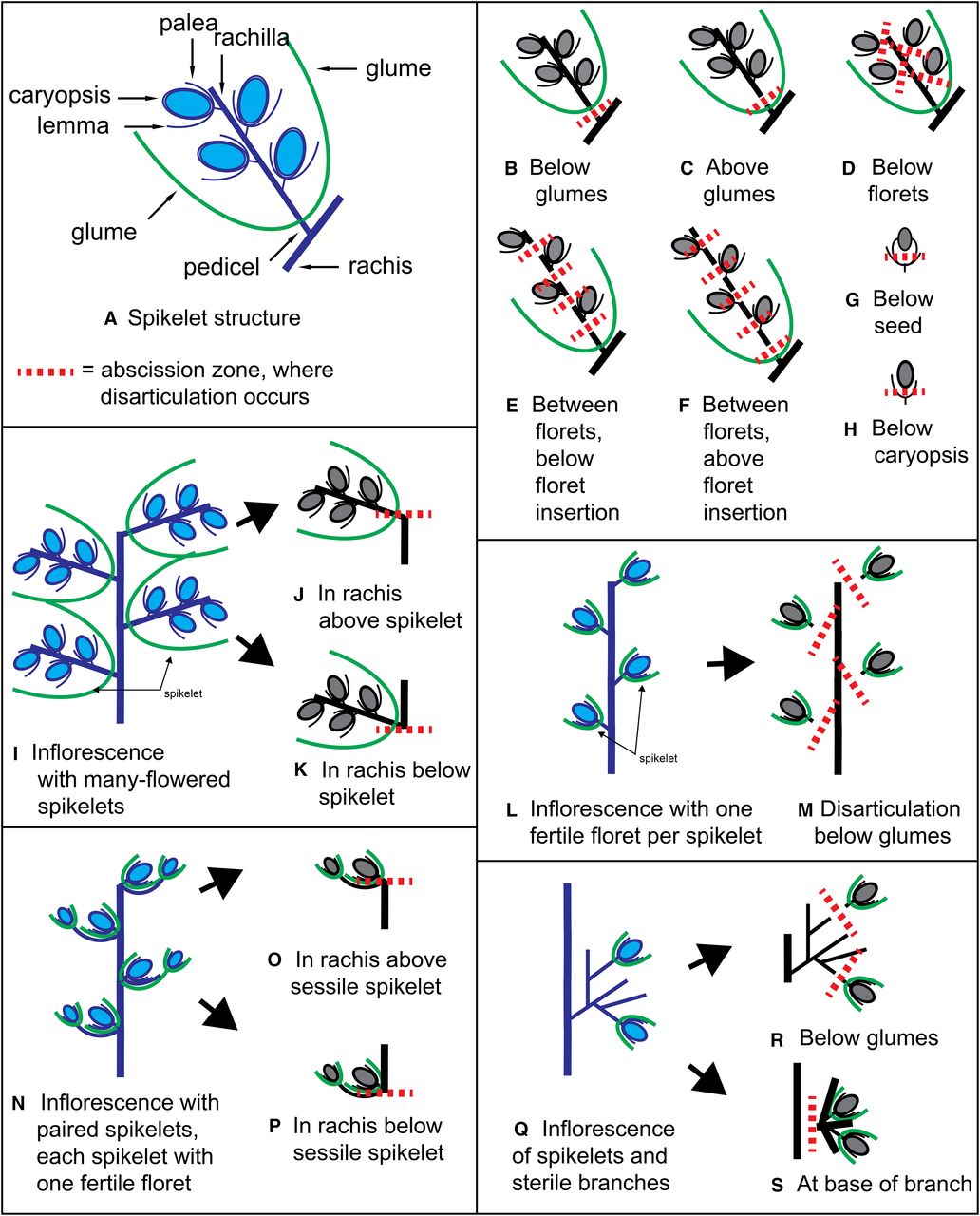
- The Nonshattering Trait in Cereal Crops – Is there a common genetic pathway that controls the shattering/nonshattering trait in cereal crops?
- Apples and Genetic Bottlenecks – Domestication generally leads to a loss of genetic variation compared to wild relatives, but apples have experienced only a mild loss. That loss may increase as commercial apple production relies on fewer and fewer cultivars.
- Improving Perennial Crops with Genomics – The nature of perennial crops can be an impediment to breeding efforts, which makes the introduction of new perennial crop varieties both time consuming and costly. Advances in genomics may help change that.
- Using Wild Relatives to Improve Crop Plants – Crop plants can be improved through the introduction of genes from wild relatives. They could potentially experience even greater improvement through systematic hybridization with wild relatives.
- Developing Perennial Grain Crops from the Ground Up – Some of the environmental issues resulting from agriculture could be addressed by switching from annual to perennial grain crops, but first they must be developed from wild species.

A small harvest of sweet potatoes (Ipomoea batatas ‘ Hong Hong’) from this year’s backyard mini-farm.
If I had to pick a favorite article in this issue it would be Think Globally, Research Locally: Paradigms and Place in Agroecological Research (Reynolds et al.). I know I said it in the post, but this article really sums up the reasons why this special issue of AJB is so important. Humans are incredibly resourceful, creative, and resilient, and as we have spread ourselves across the globe and grown our population into the billions, we have found ways to produce enormous amounts of food relatively cheaply. Frankly, the fact that anyone is going hungry or dying of starvation is shameful and appalling as there is plenty of food to go around…for now. But we are doing a lot of things wrong, and the earth is suffering because of it. If the biosphere is in trouble, we are all in trouble. Thus, we are overdue for some major shifts in the way we do things, particularly agriculture as that’s what this series of posts is all about. I advocate for science-based sustainable agriculture, and I am hopeful, thanks to this issue of AJB and other signs I’ve seen recently, that we are moving more in that direction. I’ll step off my soapbox now and leave you with an excerpt from the article by Reynolds, et al.
“There is increasing recognition that the current industrial model of agricultural intensification is unsustainable on numerous grounds. Powered by finite and nonrenewable stores of fossil fuels over the last 200 years, humans have come to see themselves, their technology, and their built environments as controllers of nature rather than interdependent with it, even as our activities threaten to exceed planetary boundaries of resilience in multiple environmental dimensions, such as climate, biodiversity, ozone, and chemical pollution. … In the ‘full world’ we now live in, continuing to use high input, highly polluting methods of food production to support continued economic growth is counterproductive to achieving food security. Continued growth of population and per capita consumption on a finite planet fails to meet the basic requirement of sustainability, that of meeting needs within the regenerative and assimilative capacity of the biosphere. And prolonging the shift to a sustainable economic paradigm risks a harder landing.”


![Grapefruit (Citrus x paradisi) - A hybrid between sweet orange (Citrus sinensis) and shaddock (Citrus maxima) that "occurred far beyond the region of domestication and rather recently [the 18th centruy]." (photo credit: wikimedia commons)](https://awkwardbotany.com/wp-content/uploads/2014/12/grapefruit.jpeg)